1 ニホンザルの分布と環境要因
岩 野 泰 三
1 はじめに
ニホンザルの分布に関する調査は,他の哺乳類に比べると例外的に古くから行なわれて来た。すなわち長谷部による1923年のアンケート調査(岩野,1974)に始まって,岸田(1953),竹下(1964),哺乳類分布調査科研グループ(1979)と最近半世紀の間に4回のニホンザルの全国的分布調査が行なわれている。しかし,これらの調査によって得られた情報は,地域によって精粗まちまちであり,全国的に一定の精度でニホンザルの分布状況を明らかにする情報を得ることが,必要とされてきた。
1978年に行なわれた第2回自然環境保全基礎調査によって得られたニホンザルの分布に関する情報は,上記の要望を満足させるものであった。また,第3次地域区画(1kmメッシュ)を単位として集計,整理されたニホンザルの分布に関する情報は他の国土数値情報との重ねあわせを可能とするので,ニホンザルの分布に影響を与える要因を分析することを容易にしている。
本報告では,これらの資料の集計,整理結果を述べると共に,ニホンザルの分布に影響を与える要因について,気候,植生,人為の面から検討を加えた。
2 方 法
昭和54年度に磁気テープに入力された第2回自然環境保全基礎調査によって得られた情報を,電算機によって集計,整理し,本報告の基礎データとした。とりまとめられたデータは,群れ生息情報,群れとは認められない少数個体の生息情報,絶滅情報,生息情報の「見た」「聞いた」の類別,年代別の情報,およびこれらの情報の第3次地域区画を単位とする区画数である。
これらの情報数と区画数を,県毎,地方毎に整理し,比較した。
次にニホンザルの分布に影響を与える要因として,気候要因としては積雪,植生要因としては亜寒帯林,人為的要因としては森林率をとりあげ,これらとニホンザルの分布に関する前記資料とを重ねあわせ,その相互関係を分析した。
さらに,ニホンザルの生息,絶滅情報を年代別にとりまとめて,分布情報の年代別変還を検討した。
3 ニホンザルの分布―第2回自然環境保全基礎調査によって得られた情報,および第3次地域区画による集計,整理の結果―
(1)生息・絶滅情報とその分布区画数
1978年に行なわれた第2回自然環境保全基礎調査の哺乳類分布調査によって得られたニホンザルに関する情報は,17075件であった。このうち,15690件(91.89%)はニホンザルの生息に関する情報であり,1385件(8.11%)はニホンザルの絶滅に関する情報であった。表1に地方別にみた生息,絶滅に関する情報数を示した。生息,絶滅両情報とも中部地方で最も多く,四国地方で最も少なかった。生息情報のうち群れの生息情報は中部地方で最も多く(2433件),群れとはみなせない少数個体の生息情報(以下少数個体生息情報と略す)は,九州地方で最も多かった(1497件)。
ニホンザルの生息,絶滅に関する情報を県別にみると,生息情報数,群れ情報数とも長野県が最も多く(それぞれ,836,658件),絶滅に関する情報は宮崎県で最も多く(158件),少数個体に関する情報は島根県で439件と最も多かった(付表1参照)。群れ情報数の多い上位5県は,長野,岐阜,山口,三重,滋賀の各県であり,少数個体情報数の多い上位5県は,島根,宮崎,大分,京都,岡山の各県であった。
ニホンザルの生息,絶滅に関する情報の位置は第3次地域区画(基準メッシュ,1kmメッシュ)を単位としてコード化され,磁気テープに入力された(環境庁,1979)。このデータから,第3次地域区画を単位とするニホンザルの生息,絶滅に関する区画数が県ごとに集計され,分布図が作製された(付表2,3,参照)。
註 北海道および沖縄にはニホンザルは自然分布しない。本報告では両道県を除いて分析を行った。
(2)生息・絶滅の情報の種類,および区画あたりの情報数
ニホンザルに関する生息情報は「見た」情報と「聞いた」情報に分けられた。表2にこれらの情報種類別の情報数を地方別に示し,あわせて伝聞情報率と第3次地域区画あたりの「見た」情報数を掲げた。
本調査における情報収集は,現地での必ずしも専門家ではない人々に対して行なわれた聞きとりによっているので,聞きとり対象者の直接観察に根拠をもたない「聞いた」情報は,「見た」情報に比べて信頼性が劣る,と考えられる。この「聞いた」情報が全情報にどの程度含まれるか(伝聞情報率)は,生息情報の信頼性を測る一つの目安である。
伝聞情報率は,東北地方で最も高く(22.23%),近畿地方で最も低かった(7.36%)。県別にみると,伝聞情報率は岩手県が最も高く(52.11%),静岡県が最も低かった*(0.77%)。ちなみに,伝聞情報率20%以上の県は岩手,宮城,山形,福島,群馬,東京,石川の1都6県であった(付表4参照)。
これらの伝聞情報率の高い県では,得られた情報によって分布図を描く際に,他県に比べて不正確になる可能性があると考えられる。そこで,第3次地域区画の1生息区画あたりの「見た」情報数が,分布図の信頼性を測る一つの規準となりうると考え,これを各県,各地方ごとに調べた**。
1生息区画あたりの「見た」情報数は,全国平均1.21(S.D.=0.08)で,各地方別に見るとほとんど差がない。県別にみると,最低値0.60(岩手県)から最高値1.63(福島県)まである程度の分散が見られた(x=1.21,S.D.=0.20)。しかしながら,この結果は,1生息区画を得るための「見た」情報数は,各県ともに1情報程度とみることができ,伝聞情報率から推測されるほど大きな相異はなかった,と言ってよいと考えられる。
*茨城県は4つの情報すべてが「見た」情報であるが,他県に比較して情報数が格段に低く,伝聞情報率の比較対象としては不適当であると考えられる。
**この場合でも「見た」情報が一地域に偏り,他地域は「聞いた」情報によって生息区画が得られ,分布図の信頼性に疑問を残すことは,論理上考え得るが,実際の情報では,そのような極端な偏りは見られなかった。
4 ニホンザルの分布に影響を及ぼす環境要因―とくに積雪,亜寒帯林,森林率との関係―
第2回自然環境保全基礎調査によって得られたニホンザルの分布図は,本種が東北地方と九州地方に疎であることを示していた(環境庁,1979)。本種の分布域がこのように特徴的な分布傾向を示すのは,東北地方ではその寒冷な気候が,九州地方では植林等の自然地域の開発が,大きな影響を与えているのではないか,と推測される。本節では,気候の指標として積雪深を,人為的要因の指標として森林率をとりあげ,本種の分布との関係を分析することとした。また,本種は亜寒帯林に適応していないとの指摘があり(上原,1977),本種の分布域と亜寒帯林の分布域との関係も,あわせて分析した。
(1)150cm以上積雪深地域とニホンザルの分布
ニホンザルは,ヒト以外の霊長類では分布の北限に位置し,深い積雪地帯に生息するため,その寒冷地適応の生理,生態的機構については従来から注目されてきた(Izawa・Nishida,1963,和田,1964,Suzuki,1965,河合他,1970,丸山他,1974,上信越ニホンザル総合調査団,1975,増井,1976,和田,1979)。しかし,ニホンザルの全国的な分布と対応させて,気候的,生態的な分布要因を分析した例はない。本稿ではニホンザルの分布域の北半,北緯35.3度以北,東経136゜以東の地域を対象として,150cm以上の積雪日数の区画とニホンザルの生息区画との関係を調べた。積雪深150cm以上の地域は,いわゆる裏日本型気候(鈴木,1962)の地域とよく一致するが,この地域内の各5kmメッシュ区画毎に150cm以上積雪深の一冬合計日数を調べ,これを10日毎に8区分しこの8区分の積雪区画とニホンザルの生息区画との関係を見た(表3)。
表3 150cm以上積雪地域におけるニホンザルの群れ生息区画数,少数個体生息区画数,絶滅区画数およびそれらの積雪地域区画数に対する割合(北緯35.3゜~42゜,東経136゜~143゜の地域内)
対象地域における150cm以上積雪地域の区画数は4453区画(ニホンザルの分布する地域の全陸地区画数の1.95%),群れ生息区画数は553区画(全国の群れ生息区画数の8.64%),少数個体生息区画数は374区画(全国の少数個体生息区画数の7.89%),絶滅区画数15,区画(全国の絶滅区画数の12.8%)であった。このことより,ニホンザルの生息区画は積雪地域により高い比率で分布していること,同時に絶滅区画の割合も高いこと,を示している。すなわち,ニホンザルの分布域は,裏日本型気候の積雪地域を特に避けるという傾向を示していない。このことは,150cm以上積雪深の地域の区画数に対する群れ生息区画数の割合(群れ生息区画率)を見れば,さらに明らかである。すなわち,積雪地域の群れ生息区画率は12.42%と全国の陸地区画数に対する群れ生息区画数の割合,2.22%をはるかに越えていた。さらに,150cm以上積雪深の日数が増加しても,その地域に対する群れ生息区画数の割合が減少しないことは注目される。これは,少数個体生息区画率にもみることができる。
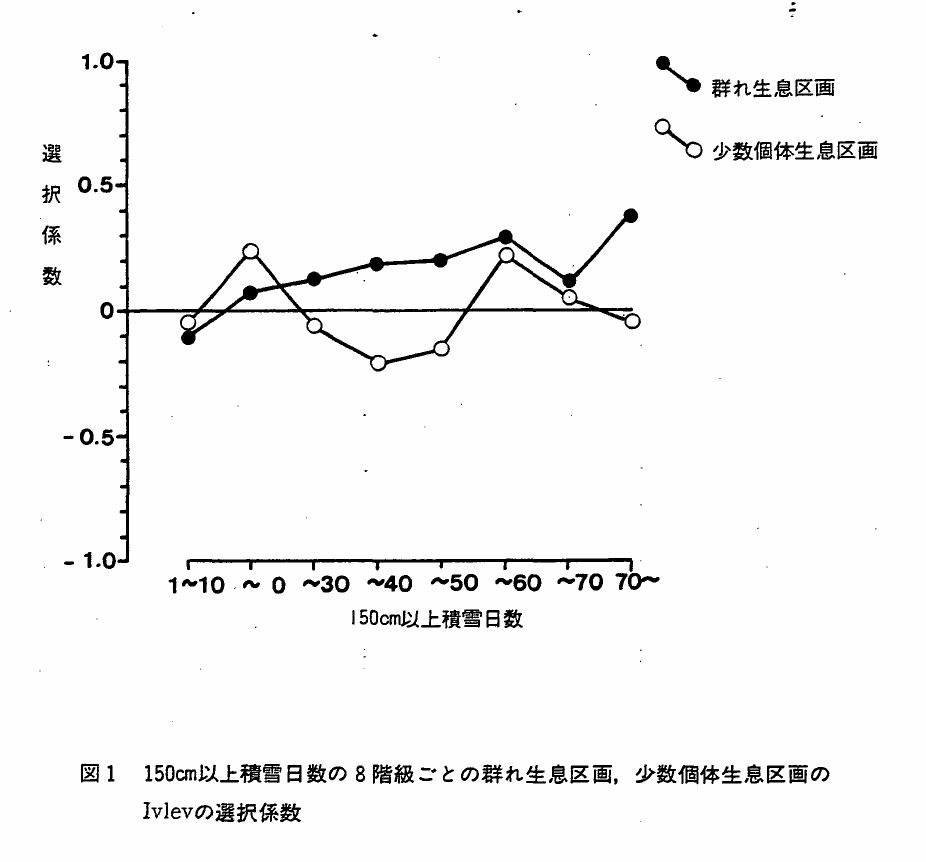
図1 150cm以上積雪日数の8階級ごとの群れ生息区画,少数個体生息区画のIvlevの選択係数
表4図1に150cm以上積雪深の地域内のどの積雪日数の階級に群れ生息区画,少数個体生息区画が集まるのを表すため,Ivlevの選択係数を示した。少数個体出現区画については,積雪日数との関係は明らかでなかったが,群れ生息区画は,積雪日数の増加と共に選択係数が徴増する傾向が見られた。
表4 150cm以上積雪深日数の8階級におけるニホンザルの生息区画
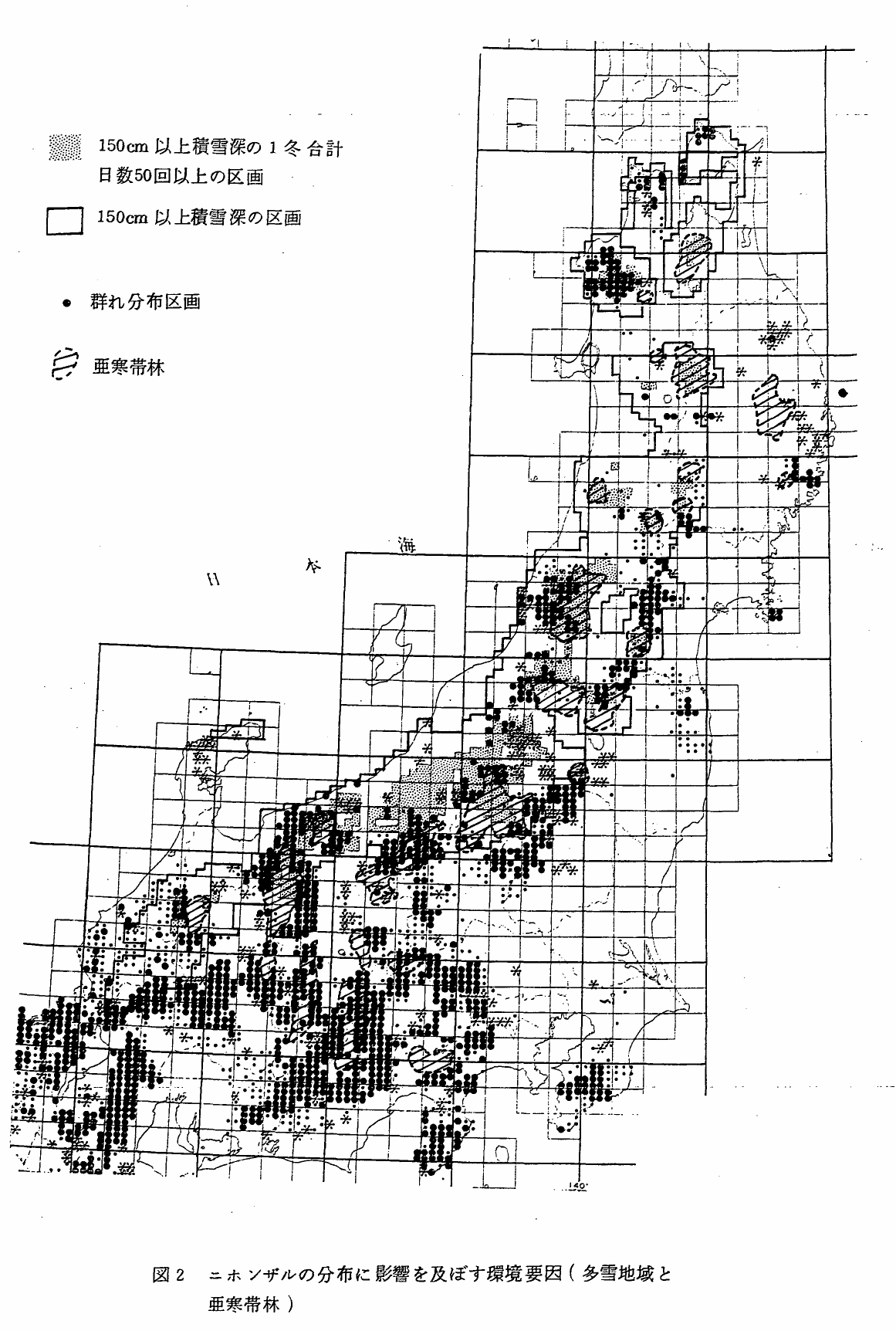
上記のニホンザルの分布域と積雪深との関係を地理的に明らかにするため,5kmメッシュで積雪深とニホンザルの分布図とを重ねあわせ,図2に示した*。ニホンザルの群れの生息域は,150cm以上積雪深が年間50日以上に達するわが国でも最も雪の深い地域,白神山地,朝日,飯豊山地,越後山地および飛騨山脈などに広く分布していた。
(2)亜寒帯林の分布とニホンザルの分布
上原(1977)は,ニホンザルの主要な食物である木本植物に注目し,亜寒帯林に生息するニホンザルでも,温帯林要素の樹種を選好するところから,ニホンザルは亜寒帯林に適応していないことを指摘した。上原はさらに進んで,ヴュルム氷期後の温暖化に伴って,亜寒帯林は収縮するが,ニホンザルがこの林を追って下北半島に達した時には,温暖化に伴う海水面の上昇のために,既に津軽海峡が成立していた。と推測し北海道にニホンザルが分布しないことの地史的,生態的説明を行った。
堀川**(1968)に加筆した上原(前掲)の図より,本州の亜寒帯林をぬき出し,ニホンザルの分布図上に重ねあわせた(図2)。これらの地域は,中部地方以北における本州の主要な山岳のほぼすべてを示している。これら,26カ所の亜寒帯林地域のうち,22カ所ではニホンザルの群れの生息区画はないか,あっても周辺のわずかな部分を占めるにすぎない。飯豊山地以北の東北地方の亜寒帯林では,朝日岳東斜面の一部を例外として,まったくニホンザルの群れは生息していない。
*図をわかりやすくするため,海岸線の1区画が海岸線に沿って連続している150cm以上積雪深1~10日間の地域は省略した。
**掘川芳雄,1968,日本の植生地図,安田女子大学紀要,2:71-75,(付図1)。
図2 ニホンザルの分布に影響を及ぼす環境要因(多雪地域と亜寒帯林)
亜寒帯林が広がっているにもかかわらず,ニホンザルの群れ生息区画がみられるのは,中部地方の4カ所である(三国山脈,金峰山,赤石山脈*,木曽山脈)。これらの地域は,三国山脈の岩菅山(2295m)周辺を除くと,すべて150cm積雪深の地域外である。
亜寒帯林の存在は,ニホンザルの分布を制限する重要な要因の一つである,特に積雪地域にあっては,ほぼ完全にニホンザルの分布を制限する,と結論できよう。
(3)森林率とニホンザルの分布
ニホンザルの分布に影響を与える人為的要因については,森林伐採による生息域攪乱の問題が1970年代より自然保護の観点からとりあげられてきた(高杉,1971,岩野,1973,村上,1975,足沢,1977,増井,1977,前川,1978,池田・江口,1978,真野目・丸山,1978,岩野,1978)。本報告では,人為的影響を測る指数として,第3次地域区画に占める森林面積の割合(以下森林率と略す)と,ニホンザルの分布域との関係を分析した。
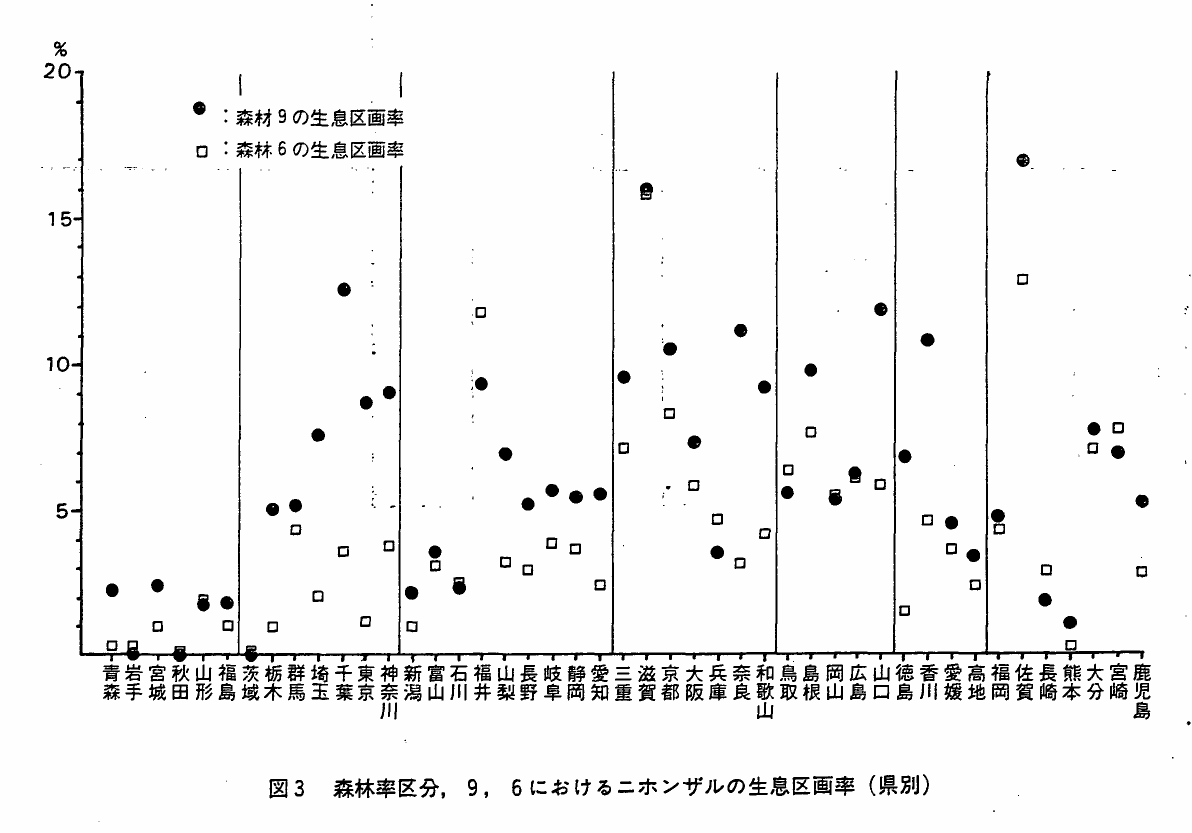
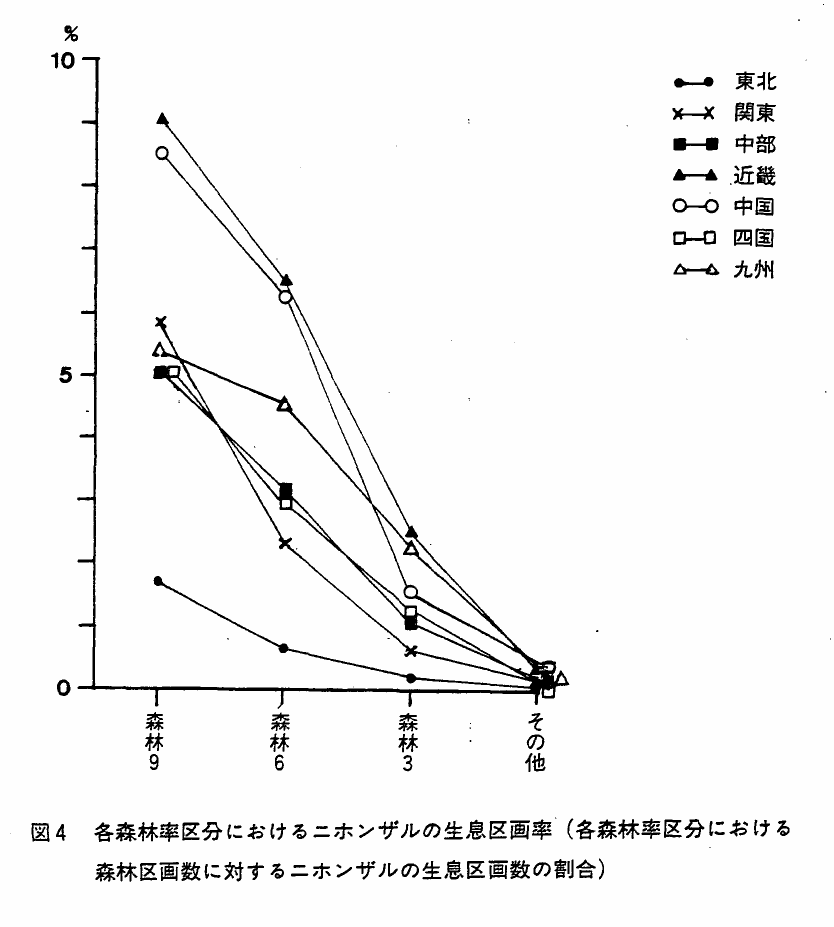
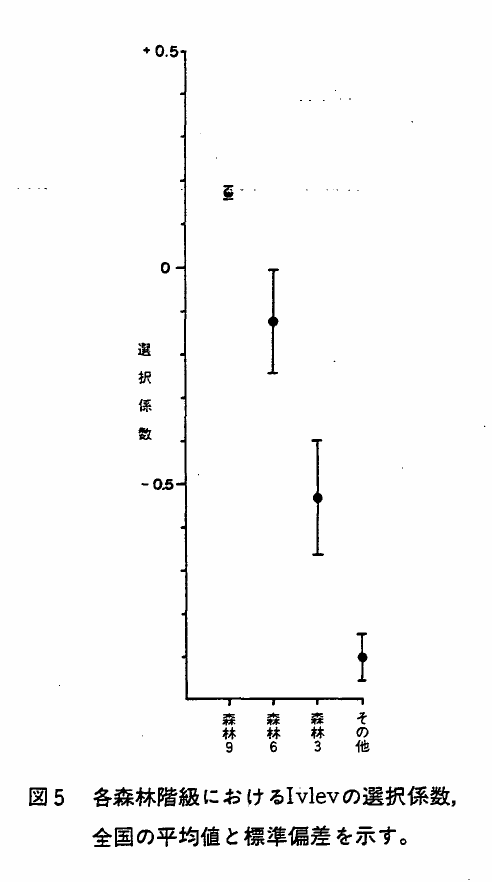
森林率を4区分し(Ⅰ章参照),それぞれにニホンザルの生息区画がいくつ含まれるかを各県別に集計し(付表6),森林率区分9と6における各県の生息区画率を図3に示した。また,各森林率区分における生息区画数とIvlevの選択係数を地方別に示し(表6),生息区画率を地方別に図4に示した。
表5 各森林率に対するニホンザルの生息区画数およびそれらの生息区画率
*もっとも,赤石山脈についての積雪の観測所がなく,実態は不明である。
図3 森林率区分,9,6におけるニホンザルの生息区画率(県別)
図4 各森林率区分におけるニホンザルの生息区画率(各森林率区分における森林区画数に対するニホンザルの生息区画数の割合)
表6 各森林率区分に対する生息区画数およびIvlevの選択係数
図5 各森林階級におけるIvlevの選択係数,全国の平均値と標準偏差を示す。
森林率区分9におけるニホンザルの生息区画数は全国で9150区画であり,森林率が下ると共に生息区画数も少なくなる傾向は,各地方,各県とも同様であった。また森林区画数に対する生息区画数の割合(生息区画率)も,一般に森林率が下ると小さくなる傾向があった。しかし,岩手,福井,兵庫,鳥取,長崎,宮崎の各県は,森林率区分9よりも森林率区分6における生息区画率の方が高く,一般的傾向を異っていた(付表5参照)。長崎県は,まったく例外的に,森林率が下るほど生息区画率が高くなった。
森林率区分9における生息区画率は,平均6.43%(s=3.96)であり,15%を越えるのは滋賀,佐賀両県のみであった。注目されるのは,東京,神奈川,大阪の各都府県で森林率区分9の生息区画率が全国平均よりも高いことである。これは大都市周辺には森林率の高い地域が少ないこと,およびその残存した少地域にニホンザルが集中して生息していることを示すと考えられる。
表7に各森林率区分におけるIvlevの選択係数*を地方別に示した。図5は,これを各森林率区分別にまとめたものである。森林率階級が下ると選択係数が下るというはっきりした傾向が見られた。しかし,同時に森林率区分6や3でもある程度の生息が見られるという点は,本種の人為的環境への適応性を物語る指数として見のがすことはできない,と考えられる。
5 ニホンザルの分布に関する情報の年代別推移
本調査では,ニホンザルの分布の年代的変遷を知る目的で,「あなたはニホンザルをどこで見ましたか」という質問項目の中に「いつごろからいますか」という欄を設け,「以前ニホンザルが生息していたところを知っていますか」という項目の中に「いつごろからいなくなりましたか」という欄を設けた。これらの設問に対する答えは,「明治,大正,戦前,昭和20年代,昭和30年代,昭和40年代,昭和50年代,不明」の8つの年代から選ぶように指定された。
表8に,「見た」および「いなくなった=絶滅」の情報数を各年代別に,各地方別に示した。「見た」情報総数13977件のうち,最も多かったのは,昭和40年代4452件(31.9%)であり,昭和50年代の情報は279件(2%)と最も少なかった,「絶滅」情報総数は1521件で,昭和40年代が361件(23.7%)と,やはり最も多く,昭和50年代は25件(1.6%)と最も少なかった。
昭和50年代の情報件数は,全年代中最も少ないが,これは昭和50年より調査時点(昭和53年)まで4年間におけるニホンザルの出現や絶滅というニホンザルの分布域の変動に関する情報として貴重なものであるといえよう。すなわち,調査時点(昭和53年)まで最近4年間にニホンザルの分布域の約2%が(情報数が分布域の関数であると仮定すれば)変動した,と言えよう。
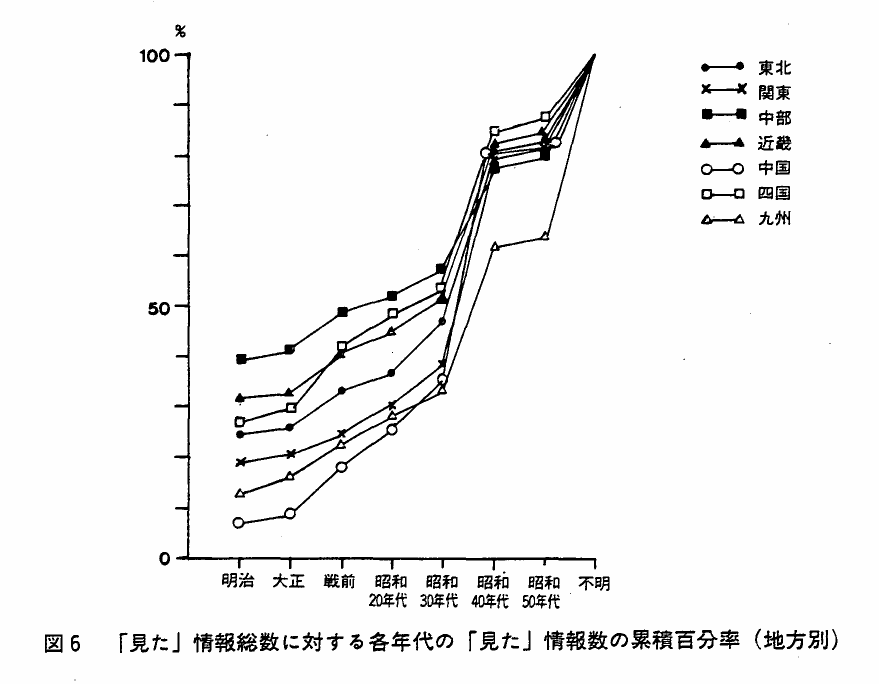
各年代の「見た」情報数の「見た」情報総数に占める割合を見ると,九州地方を除く各地方とも,時代不明はほぼ20%であった。また明治時代の情報割合は,中野,近畿地方に高く,中国,九州地方に低いという傾向が見られた(図6)。すなわち,古くから存続していることが知られている分布域は,中部,近畿地方に比べると中国,九州地方では少なかった。もっとも,時代不明の情報が九州地方では多いので,この傾向が,両地方での最近になってからの分布域の拡大を示す,とは結論できない。
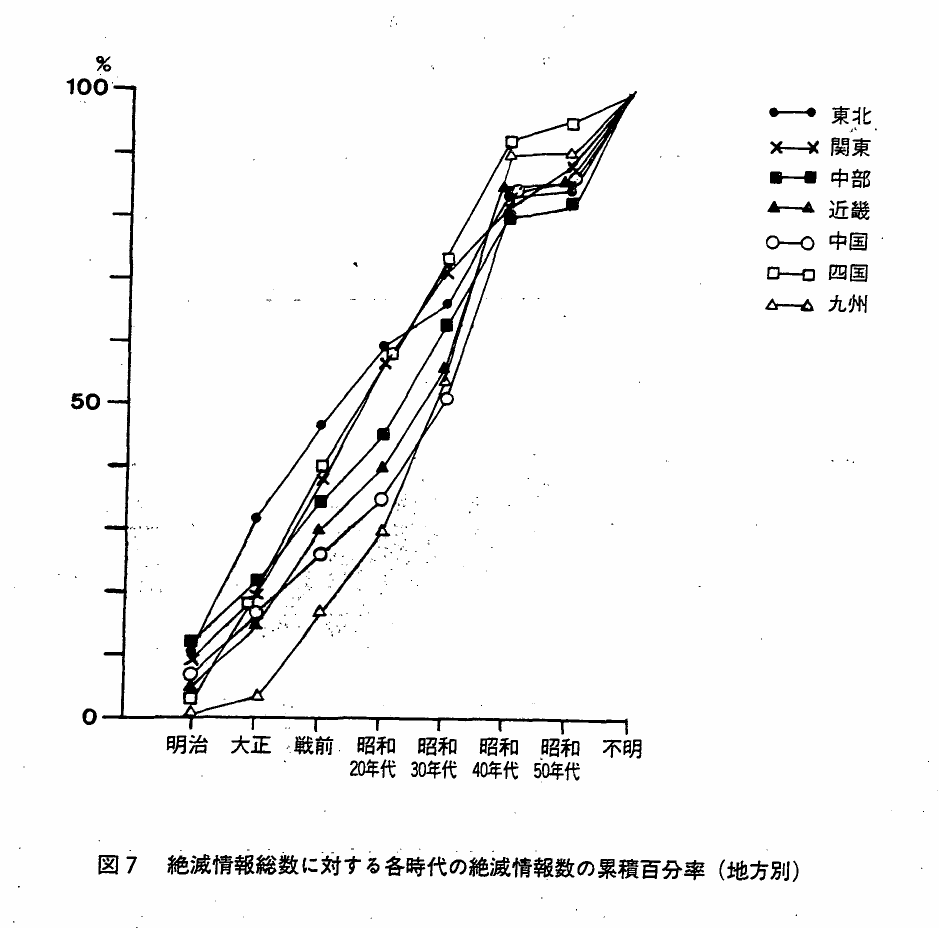
「見た」情報は,各地方とも昭和30年代まで同じような割合で増加し,昭和40年代に突然ふえる,というまったく似かよった年代分布を示すが,絶滅情報はこのような年代的に特徴のある型を各地方とも示さなかった(図7)。
ニホンザルの分布域の年代的変遷を知る目的で設定された本項目は,問いがやや抽象的であったこと,被調査の記憶だけに頼っていること,のために,情報としては信頼性が少ない。絶滅情報と比較すると「見た」情報は年代的変化の一定のパターンがあった,と言えるが,これが昭和40年代における分布域の拡大を意味するか,どうかについてはこれを客観的に調べる資料がない。分布域の変動については,本調査と同様の調査を繰りかえし,アンケートによる情報の,ことに年代に関する信頼性の程度をみきわめる必要があると考えられる。
図6 「見た」情報総数に対する各年代の「見た」情報数の累積百分率(地方別)
図7 絶滅情報総数に対する各時代の絶滅情報数の累積百分率(地方別)
6 考 察
(1)ニホンザルの分布を制限する環境要因
本分析の結果,最も注目されるのは1ニホンザルの分布域が150cm以上の積雪深が年間50日以上みられるという多雪地域を避けない,という傾向を見せたこと,および,2ニホンザルの分布域は亜寒帯林の存在によって制限されること,の二点である。
本調査のアンケート項目にはニホンザルが見られた時期を記載する欄がないので実際には,サルは冬期には深い積雪を避けてより低い山麓地域に移動しているのだが,本調査の情報では夏期の生息域が反映されて,上記のように深い積雪地域をニホンザルの群れは選んで生息するという結果になったのではないか,と疑うことはできる。丸山他(1974)は,ニホンザルの群れが,積雪期に低標高地域へ季節的に移動することを報告している。しかし,長野県志賀高原でのニホンザルの野生群の周年的な追跡によれば(和田,前掲),季節的移動は遊動域の全体をかえるほど大きくはない。すなわち,ニホンザルの遊動域の季節的変化は,5kmメッシュの枠を大幅にはみ出すほど大きな移動ではない。したがって,本調査結果は,多雪地帯においても,ほぼ正確にニホンザルの分布の現状を反映していると考えられる。
ニホンザルの分布域が多雪地域に広く見られる理由として,(イ)ニホンザルは 生理的,生態的に積雪に適応していること,(ロ)多雪地域は造林に適さないので,これらの地域は自然林として残る可能性が高く,結果的にニホンザルの生息地を保全したこと,(ハ)ニホンザルの群れの生息区画のある多雪地域はいずれも大きな山塊であり,造林,狩猟等の人為的影響をうけにくい地域であること,などが考えられる。
では,何故東北・北陸地方の大山塊の多雪地帯で,ニホンザルの生息しない地域が見られるのであろうか?この問いに対する答えは,三通りである。
(イ)人為的要因,ことに,ニホンザルに対する集団的な狩猟の有無。裏日本では,冬季にサル狩を村民総出で行なってきた地域もあり,その影響は見のがせない。しかし,過去半世紀の狩猟統計によれば,東北,北陸地方のニホンザルの狩猟数は,他地方に比較すればむしろ少なく,特にニホンザルが狩猟鳥獣としては認められなくなった戦後では,これらの地方ではニホンザルはほとんど猟獲されていない(渡辺,1978)。したがって,多雪地域でのニホンザルの分布の狩猟による消滅は,戦前の,それもかなり古い時代のものに限られると考えられる。
(ロ)地形的要因
気象観測上,同じ積雪深であっても,平らな地形と急峻な地形では,積雪のニホンザルに与える影響は異なる,と考えられる。平担地での深い積雪は,ニホンザルの長距離移動を妨げ,採食を困難し,イヌや人間の捕食,狩猟圧を高める。しかし,急峻な崖を多く含む地形では,ニホンザルは安全な逃げ場と採食地を持つことができる。
八甲田山,八幡平,朝日山地,尾瀬ケ原,魚沼・頸城丘陵などのニホンザルの分布しない多雪地域の地形がなだらかであることは,上記の考察を支持するものであると考えられる。
(ハ)植生上の要因
上原(前掲)の指摘する通り,ニホンザルは亜寒帯林要素の木本植物を,その主要食物のなかにとり入れていない。また,丸山ら(前掲)の観察は,積雪地を利用しない,というよりも積雪期においては亜寒帯林を利用しないことを示していると考えられる。したがって中部地方以北の山岳地帯上部の亜寒帯林はそれが広い場合は,ニホンザルの分布を制限する要因となっていることが予想される。青森県の二つの多雪地域,白神山地と八甲田山を比較すると,八甲田山の方が亜寒帯林の分布域が広い。そして,白神山地にはニホンザルの群れの広い生息域があり,八甲田山には,まったくサルは生息していないのである。ニホンザルの分布と積雪,植生要因の関係を以下のようにまとめることができる。
(1)多雪地域(150cm積雪深日数50日以上の地域)はニホンザルの分布に直接の制限を与えない。しかし,多雪地域で亜寒帯林がある場合は,ニホンザルの分布は制限される(例外,朝日岳東斜面,岩菅山周辺)。
(2)亜寒帯林は明らかにニホンザルの分布の制限要因の一つであって,深い積雪がみられない地域でもニホンザルの分布を制限する(例:北上山地,富士山,八ケ岳,御嶽山*)。
(3)深い積雪のない地域では,亜寒帯林でもニホンザルの分布が見られる(例:赤石山脈,木曽山脈,金峰山)。これは深い積雪がニホンザルの分布を制限する要因の一つであるという逆の示唆というよりも,これらの地域の他の環境要因(たとえば,亜寒帯林といっても北陸,東北地方のものに比べると,その種組成が異なるといった植生上の,あるいは気温の高さなどの気候上の要因)の多雪地域との相異を示すものと考えられる。
森林率に示される人為的影響は,ニホンザルの分布を制限する最大の要因である。ニホンザルは森林率の低い区画(森林3)でも見られるが,それらの地域での生息率はきわめて低く,このような環境は,ニホンザルの分布を確実に制限している。この点の分析を進めるためには,森林の組成(自然林と植林の区別),道路密度,人口密度などの要因とのニホンザルの分布情報の重ねあわせが必要である。
*北上山地を除くこれら三山はいずれも高標高の火山であるという点は注目される。
(2)ニホンザルの保護
ニホンザルの地域個体群の大きさを示す目安となるのは,群れ生息区画率(陸地区画数に対する群れ生息区画数の割合)である,と考えられる。群れ生息区画率は東北地方で最も低く,九州地方がこれについで低い。両地方では,群れ生息区画数も少ないので,ニホンザルの保護上留意すべき地方として考えられる。ことに東北地方では,人為の影響を受けにくい高山が,ニホンザルの分布を制限する亜寒帯林となっているため,ニホンザルの存続は常に危険にさらされていると言えよう。東北地方のニホンザルの分布域は,単に霊長類の分布の北限というばかりでなく,その生態的適応の極限を示すものとして,特に重要な位置を占めるものと考えられる。東北地方におけるニホンザルの分布域の保全は,本種の保護にとって重要である。
7 引用文献
足沢貞成,1977,下北のニホンザル―その現況と変遷―,にほんざる,3:11-22.
哺乳類分布調査科研グループ(文責:古林賢恒,岩野泰三,丸山直樹),1979,カモシカ・シカ・ヒグマ・ツキノワグマ・ニホンザル・イノシシの全国的生息分布ならびに被害分布,生物科学,31(2):96-112.
池田啓・江口和洋,1978,九州北部におけるニホンザルの分布,にほんざる,4:49-60.
Ivlev,V.S.,1955(児玉康雄,吉原友吉訳,1965),魚類の栄養生態学,新科学文献刊行会,米子,PP.261
岩野泰三,1973,追いつめられた房総のサル,アニマ,1(8):31-35.
岩野泰三,1974,ニホンザルの分布,にほんざる,1:5-62.
岩野泰三,1978,ニホンザルの危機と保護,にほんざる,4:122-131.
Izawa,K.and T.Nishida,1963,Monkeys living in the northern limits of their distrabution,Primates,4(2):66-88.
上信越ニホンザル総合調査団,1975,多雪地域のニホンザル―志賀A群の総合調査,生理・生態,16:1-107.
環境庁,1975,第2回自然環境保全基礎調査動物分布調査報告書(哺乳類)全国版,pp.152.
河合雅雄他,1970,白山周辺におけるニホンザルの生態学的調査Ⅰ,日本自然保護協会中部支部,白山学術調査団編,白山の自然,335-343,石川県.
岸田久吉,1953,代表的林棲哺乳動物ホンザル調査報告,鳥獣調査報告第14号,農林省林野庁,pp.92+pl.Ⅲ.
前川愼吾,1978,和歌山県におけるニホンザルの分布状況について,にほんざる,4:14-31.
真野目博,丸山直樹,1978,ニホンザルの保護と農林業,にほんざる,4:112-131.
丸山直樹,常田邦彦,遠竹行俊,岡林良一,1974,裏男体有料道路がシカ,サルの生態におよぼす影響,日光の自然を守る会会報,大自然12:7-15.
増井憲一,1976,ニホグザルの群れの大きさと構成についての記録―群れの大きさと個体群に関するこれまでの見解の検討―,生理生態,17:185-194.
増井憲一,1977,白山地域のニホンザルの分布と保護上の問題点,にほんざる,3:80-87.
村上興正,1975,生態系の保護と管理Ⅱ―動物―,共立出版,東京,pp.158.
鈴木秀夫,1962,日本の気候区分,地理学評論,35(5):205-211.
Suzuki,A.,1965,An ecological study of Japanese monkeys in snowy areas;Focused on their food habits,Primates,6(1):31-72.
高杉欣一,1971,われわれにとっての自然保護,房総スカイライン問題の渦中から,自然,26(12):90-95.
竹下完,1964,野生ニホンザルの分布及びポピュレーション(上)―アンケート調査による―野猿,19:6-14.
竹下完,1964,野生ニホンザルの分布及びポピュレーション(下)―アンケート調査による―野猿,20-21:12-21.
上原重男,1977,食性からみたニホンザルの適応に関する生物地理学的研究,加藤泰安,中尾佐助,梅
棹忠夫編,形質・進化・霊長類,今西錦司博士古稀記念論文集,187-232.
和田一雄,1979,野生ニホンザルの世界一志賀高原を中心とした生態一,講談社,東京,pp.258.
渡辺隆一,1978,狩猟統計及び鳥獣関係統計からみたニホンザルの捕獲について,にほんざる,4:91-95.
8 要 約
1 ニホンザルの分布に関する第2回自然環境保全基礎調査の情報(合計17075件)を集計し,第3次地域区画を単位として,これらを整理し,図化した。
2 第3次地域区画を単位とするニホンザルの生息区画総数は11438区画,絶滅区画総数は1180区画,群れ生息区画総数は6397区画,少数個体生息区画総数は4738区画であった。
3 陸地区画数に対する群れ生息区画数の割合(群れ生息区画率)および,陸地区画数に対する少数個体生息区画数の割合(少数個体生息区画率)は,共に東北,北陸地方で低い値を示した。
4 ニホンザルの分布に影響を及ぼす環境要因として,1.5m以上の積雪,亜寒帯林植生および陸地面積あたりの森林面積(森林率)をとりあげた。これらの要因とニホンザルの分布域との関係を分析した結果(1)ニホンザルは,1.5m以上の積雪の日数が50日以上の多雪地帯にも他地域とかわりなく分布する,(2)亜寒帯林の存在は,ニホンザルの分布を制限する,(3)森林率の高い地域をニホンザルは選好する傾向がある,ことが明らかになった。
5 ニホンザルの生息情報は各地方とも,昭和40年代に最も多く,明治時代がこれに次いだ。絶滅情報数は,時代的偏りはみられなかった。
Summary
1. 17,075 items of informaion concerning the distribution of Japanese monkeys, obtained by the Second Basic Survey of the Natural Environment, were analyzed and synthetized. The distribution maps based on this data were drawn to the scale of the third national grid map (1 km2 grids).
2. The total number of occupation grid squares is 11,438, extinction grid squares 1,180, and the grid squares occupied by monkey troops 4,738.
3. The relationship between monkey distribution patterns and environmental factors was analysed and the following results obtained:
(1) Japanese monkeys, found in deep snow-covered areas (1,5 m of snow-fall lasting more than 50 days), are found equally in the shallower snow-covered areas.
(2) The major factor limiting monkey distribution is sub-alpine forests.
(3) Monkeys seem to prefer forest areas with a coverage-rate of 70 per cent or more.
4. Some tendencies concerning the periods of greatest frequency and extinction for monkey populations were examined. Data on monkeys in Japan is richest for the years around 1965. The next richest period for data is the Meiji era Data on monkey extinction shows no such tendency.
付表2 ニホンザルの生息・絶滅区画数およびそれらの全区画数に対する割合
付表3 ニホンザルの群れ・少数個体生息区画数とそれらの全区画数に対する割合
目次へ
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}