5 イノシシの地理的分布とその要因
常田 邦彦・丸山 直樹
本報告は,「第2回自然環境保全基礎調査」の一環として実施されたイノシシ(Sus scrofa)の分布調査の結果にもとづき,本種の地理的分布とその要因分析を試みたものである。この調査による本種の地理的分布については,「第2回自然環境保全基礎調査,動物分布調査報告書(哺乳類),全国版」(1979)で報告したので参照されたい。本報ではこれにもとづき,さらに地理的分布構造からの分析を試みることにした。
ところで,本種の地理的分布要因についてはいくつかの報告が行われているが(千葉,1959,1964;朝日,1975;高橋,朝日,1978;鳥居,1978;など),多くは地域的分析にとどまっており,一定の方法にもとづき,短期間に全国的に集められた資料を使っての分析は,本報が初の試みと考えられる。
1 本種の分布と積雪
千葉(1963)は,本種の分布限界が最深積雪1mラインにほぼ一致すると述べている。しかし,一般に有蹄類は後肢附関節の高さ以上の積雪によって行動を著るしく阻害され(Gilbert et al,1970),本種では約30cmの積雪深が行動阻害につながると考えられている(朝日ら,1975)ので,ここでは積雪深30cm以上の日数をとりあげることにした。
このため,気象庁発行「都道府県気象月報」にもとづき,1968~1978年までの冬期10シーズンについて,観測地点毎に積雪深30cm以上平均日数を求めた。但し,6日以上の欠測日があるシーズンおよび6シーズン以上の欠測がある観測地点は除外した。この結果を地図上に落し,地形・高度を考慮しながら等積雪日数線を引き,5kmメッシュによる30cm以上積雪日数分布図を作成した。
表1 30cm以上積雪日数区分毎のイノシシの生息区画数と絶滅区画数
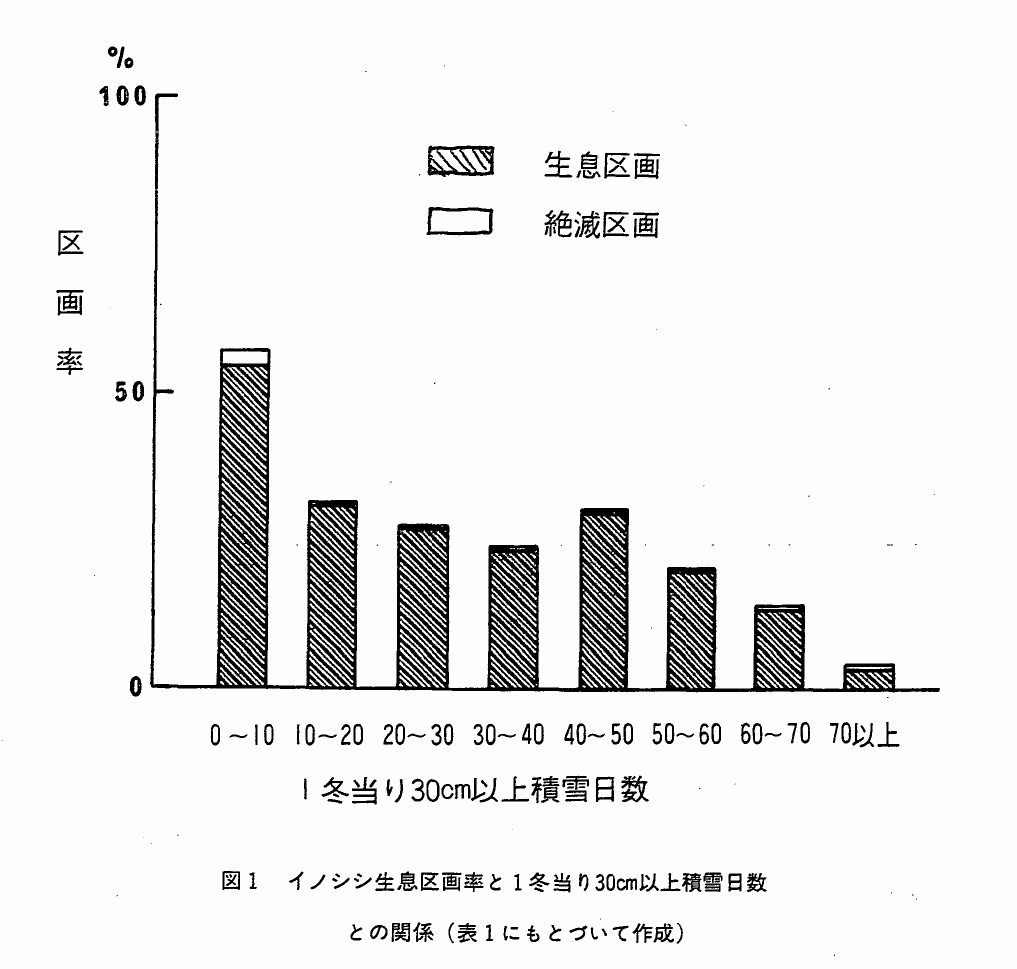
図1 イノシシ生息区画率と1冬当り30cm以上積雪日数との関係(表1にもとづいて作成)
表1にもとづく図1は,1冬当り30cm以上平均積雪日数区分に対応した区画総数に対する生息区画率を示している。積雪日数が10日未満では全区画の50%以上に生息が認められるが,積雪日数が多くなるにつれて生息区画率は減少し,積雪日数区分70日以上ではわずかに3.3%(56区画)を示すにすぎない。この56区画の位置と内訳は,長野県1区画,岐阜・石川・福井の県境地帯51区画,兵庫県4区画となっている。
本種の積雪に対する自然分布の限界は,1冬当り30cm以上平均積雪日数70日を一応の目安とすることが可能と考えられるが,なおこの線を越える例外的な生息区画が認められる背景としては次のことが考えられる。(1)本種は実際には,この限界を越えて生息が可能である。(2)本種の分布は実際にはこの限界内におさまるのだが,積雪日数分布の表示上の誤差が影響している。(3)Sokolov(1962)や朝日(1975)が指摘している本種の季節的移動による夏と冬の生息地を区分しなかったため。
本種の生息が,青森・秋田・山形・新潟・富山の各県をなじめとした多雪地帯で認められないのは,まさしく自然的分布制限要因としての積雪が主に原因しているためであろう。
2 本種の分布と森林率
表2は,森林率区分に対応した区画総数と生息区画数である。
全国合計をみると,本種の全生息区画の約80%(16,814区画)が,森林率70%以上の「森林9」に含まれ,森林率が低い区分ほど生息区画数が少なくなっている。森林率10%未満の区分である「その他」ではわずか0.6%,124区画に生息が認められるに過ぎない。
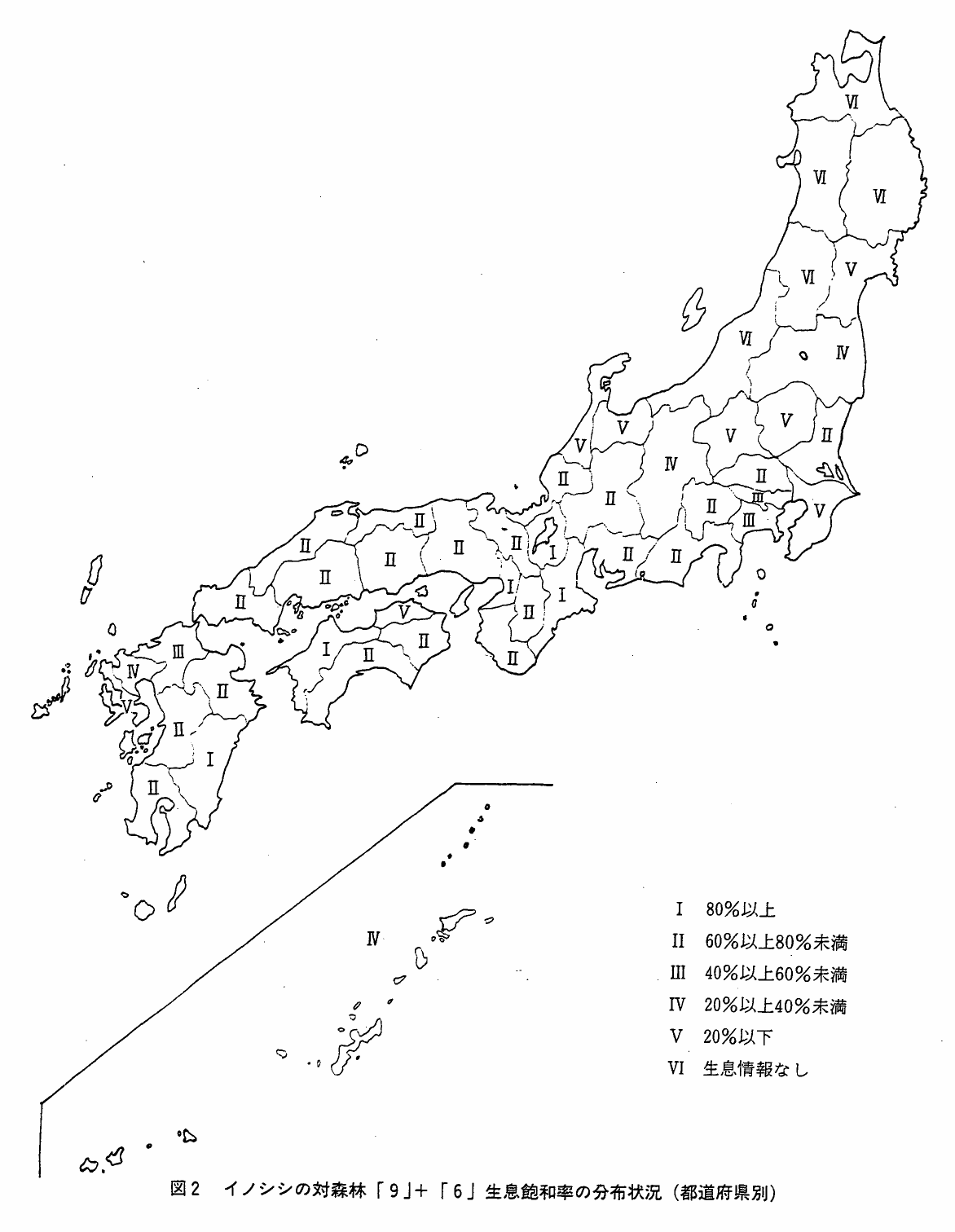
図2 イノシシの対森林「9」+「6」生息飽和率の分布状況(都道府県別)
地方別にみても,「森林9」に含まれる生息区画の割合は,東北地方と九州地方で60%とやや低いほかは,いずれも80%以上を示し,森林率が低くなるほど生息区画数が少なくなる傾向は一致している。
県別にみてもこの傾向は明確に認められた(付表3)。福島,福井,三重,宮崎の4県では「森林9」よりも「森林6」の生息区画数が多かったが,両区分に含まれる生息区画数は,全生息区画の90%以上に達している。
以上の結果から,本種は明らかに森林地帯を中心に分布しているといえる。
ところで朝日ら(1975)と鳥居(1978)は,本種の生息環境として,森林・耕作地・人家が混在する場所をあげているが,これは本報の分析と矛盾するものではない。今回の分析からも,本種は確かに耕作地や市街地がある程度存在する地域にも生息することがわかる。だが一方でその比率が高くなると殆んど生息しなくなることも事実である。
表3は,森林区分「9」「6」に含まれる区画数に対する生息区画率すなわち生息飽和率をみたものである。
生息飽和率の平均は北海道を除いて計算すると49.6%で,まだまだ分布域を拡大し得る余地はありそうだが,すでにみたように,北陸・東北地方は森林の状態がよくても多雪のために生息が難かしいから,多雪地帯を除くと飽和率はさらに高くなる。関東地方は42.5%とやや低いが,新潟・富山・石川・長野4県を除いた中部地方が66.8%,四国地方58.8%,九州地方52.9%,中国地方68.5%と全国平均よりも高く,近畿地方は82.0%ときわめて高い値を示している。
生息飽和率80%以上の飽和率区分Ⅰの該当地域は5府県で,近畿地方が大阪・三重・滋賀の3府県,四国地方では愛媛県,九州地方では宮崎県となっている。次いで生息飽和率が60%以上80%未満の区分Ⅱを示すのは21府県で,近畿・中国地方を中心にして九州・四国・中部および関東地方の一部に分布している。飽和率40%以下の区分は,先述した多雪地帯である北陸・東北地方に分布するとともに,四国の香川県,九州の佐賀・長崎・沖縄の各県となっている(図2)。
多雪地帯以外の地域で生息飽和率を下げている原因としては後述する狩猟による捕獲が第一に考えられる。
多雪地帯を除けば,この飽和率の低い地域ほど今後さらに分布を拡大し得るわけであるが,ここでいう「森林」には本種にとって必らずしも好適でないものも含まれているから注意を要する。
3 本種の出現情報とその推移
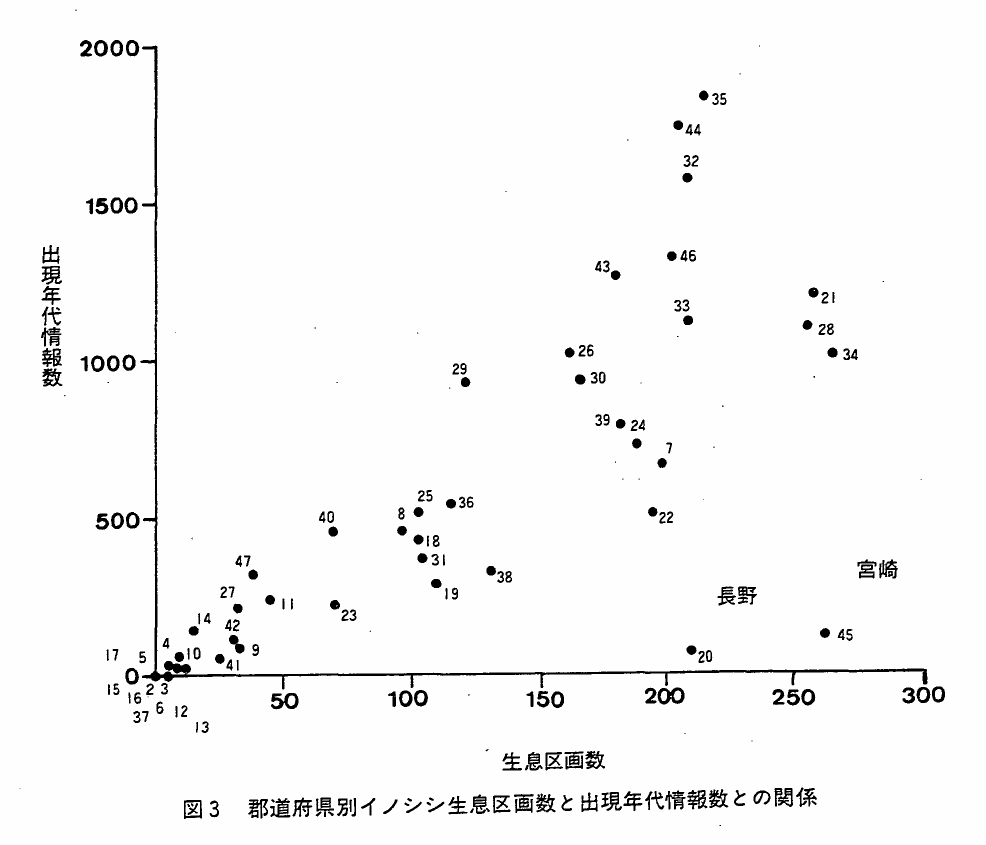
図3は,都府県別の本種の生息区画数と出現年代情報数との関係をみたものである。両者には正の相関が認められ,生息区画数が多くなるほど,出現年代情報数が多くなる傾向が認められる。当然のことながら,これは地元民と本種との関係量が生息域の拡がりと一定の関係を有していることによるものである。但し,長野・宮崎両県の情報数が他県と比べてきわめて少なくなっているが理由は不明である。
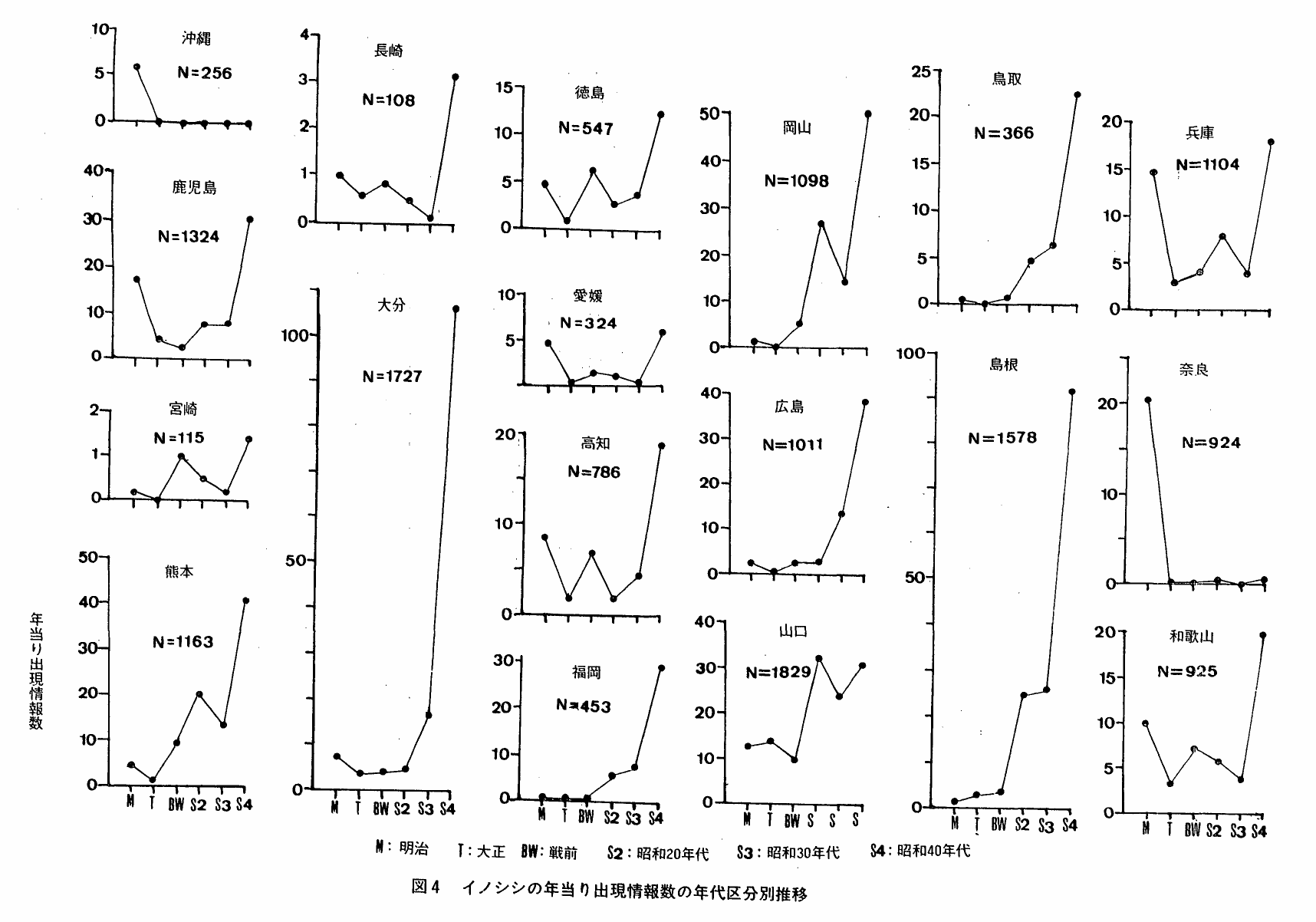
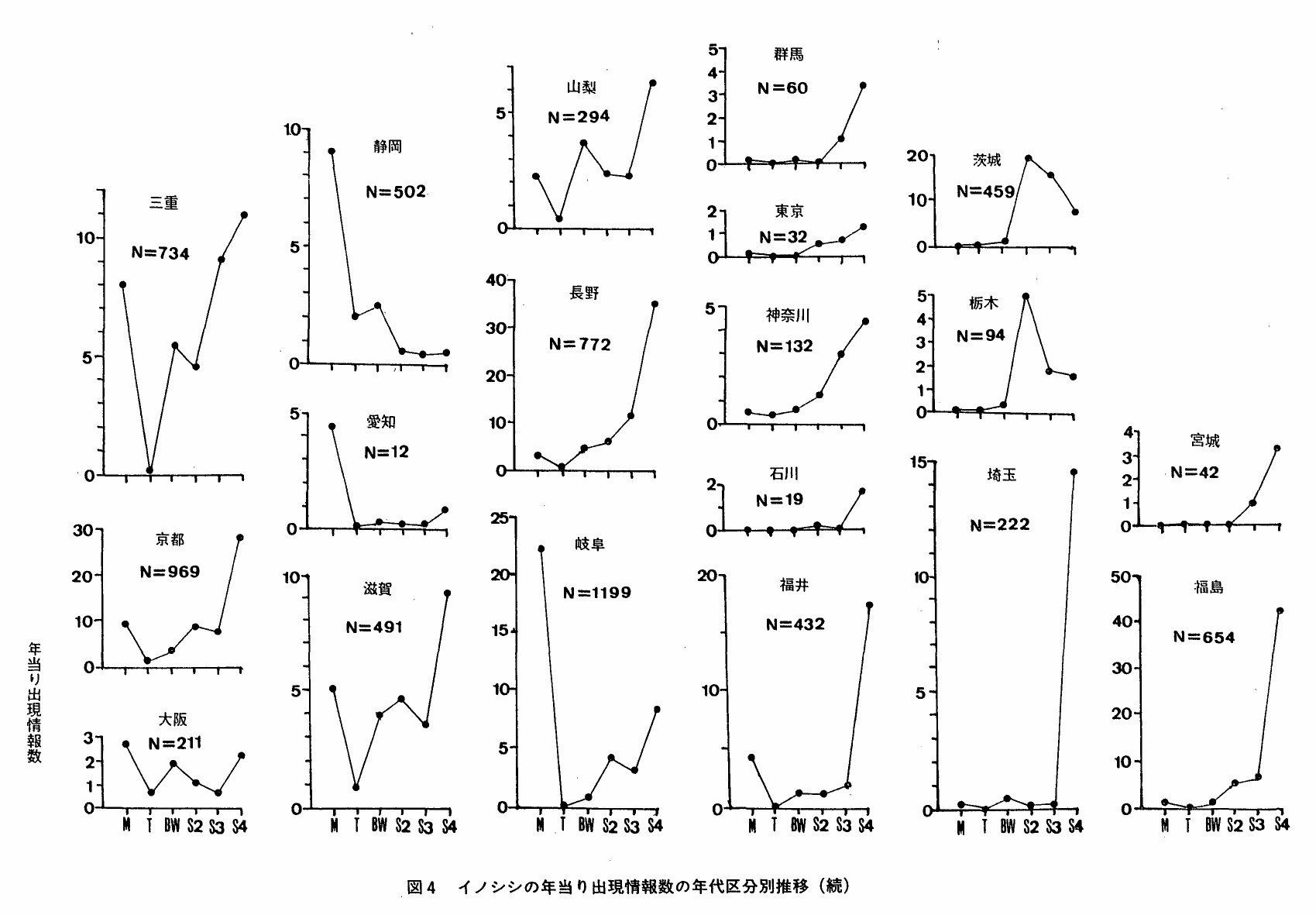
図4は,本種の年当り出現情報数の年代区分別推移をみたものである。都府県によって様々なパターンが認められるが,おおよそ次の5種類に区分することが出来る。
図5 イノシシ年当り出現惰報数推移パターンの分布状況(都道府県別)
J字型:年当り出現情報数は,明治・大正は殆んどないかわずかで,時代が進むにつれて多くなる。
このパターンは,情報数の増加開始時期から次のように整理できる。
昭和40年代;石川・埼玉・福井
昭和30年代;群馬・宮崎・広島・大分
昭和20年代;福島・神奈川・東京・島取・島根・福岡
戦 前;長野
N字型:年当り出現情報数の推移は,増加→減少→増加を示す。山口・岡山・熊本(昭和20年代に最初のピークが現われる),宮崎(昭和戦前期に最初のピークが現われる)
W字型:年当り出現情報数の推移は,滅少→増加→減少→増加を示す。山梨・滋賀・兵庫・大阪・和歌山・高知・徳島
V・U字型:年当り出現情報数は減少次いで増加に移行する。岐阜・京都・三重 年当り出現情報数は,明治以後減少し,しばらく横這状態を続けた後,再び増加を示す。愛媛・鹿児島
L字型:年当り生息情報出況数は,明治と大正で減少し,以後横這を示す。沖縄・愛知・静岡・奈良
∧ 型:年当り生息情報出現数は,昭和20年代に増加し,以後減少を示す。栃木・茨城
図5は,上述した類型の分布状況をみたものである。J字型は,主に東日本,北陸,山陰,北九州地方に分布しており,一方,VU,W,N型は南九州,四国,近畿地方といった西南日本を中心に分布している。A型は北関東,L型は東海地方,奈良県,沖縄県となっている。
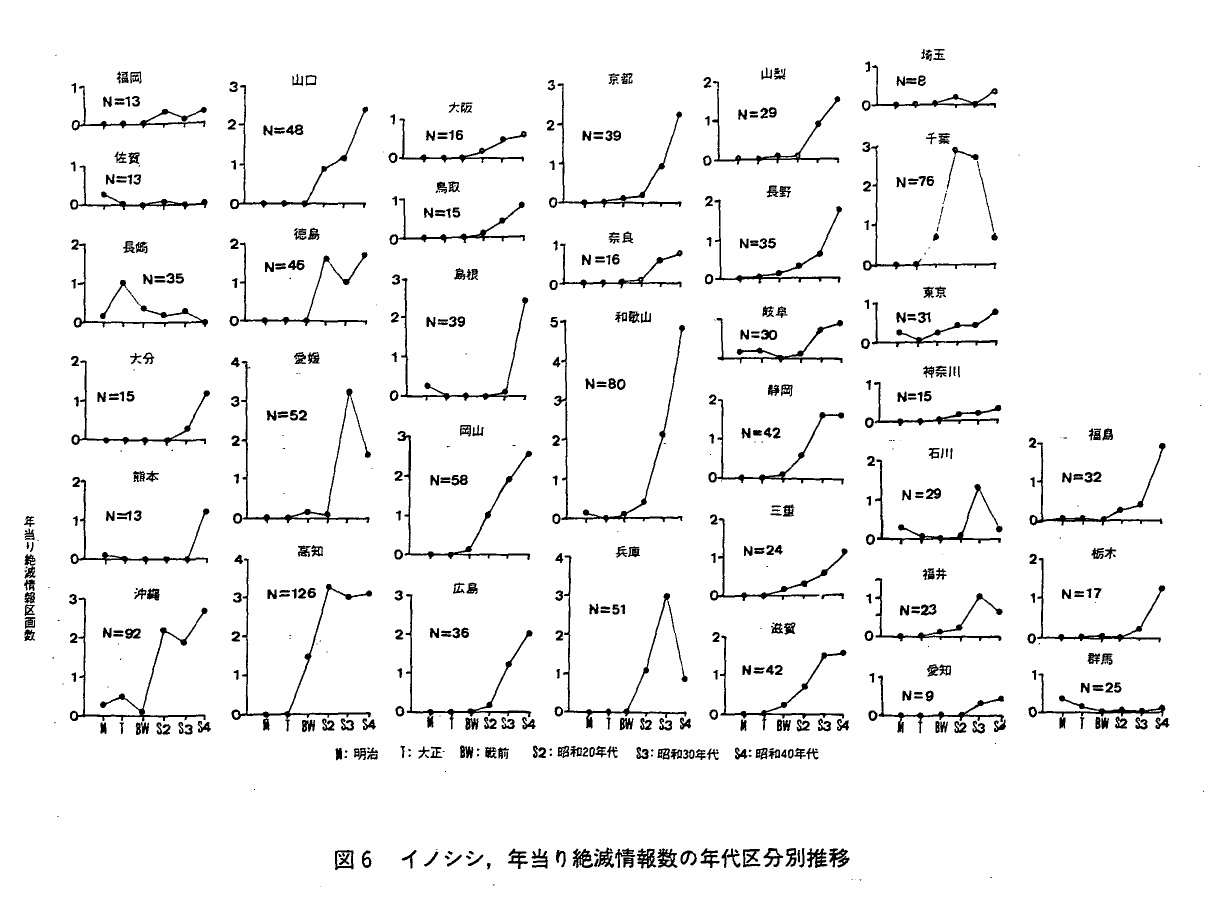
4 本種の絶滅情報数の推移
図6は,年当り絶滅情報数の年代区分別推移をみたものである。都府県によって種々なパターンが認められるが,おおよそ次の4種類に区分することが出来る。
S字型:年当り絶滅情報数は明治,大正は少ないが以後増加し,振動または横這を示す。高知・沖縄・徳島・静岡・滋賀・埼玉・福岡・愛知・岐阜・大阪・奈良
J字型:年当り絶滅情報数は明治,大正は少ないが以後J字状に増加する。鳥取・岡山・広島・山口・和歌山・大分・熊本・島根・三重・京都・東京・山梨・長野・福島・栃木・神奈川
∧ 型:年当り絶滅情報数は明治・大正は少ないが以後増加し,現在はやや減少している。愛媛・福井・兵庫・千葉・石川
D 型:年当り絶滅情報数は明治,大正に多く以後減少傾向を示す。長崎・佐賀・群馬
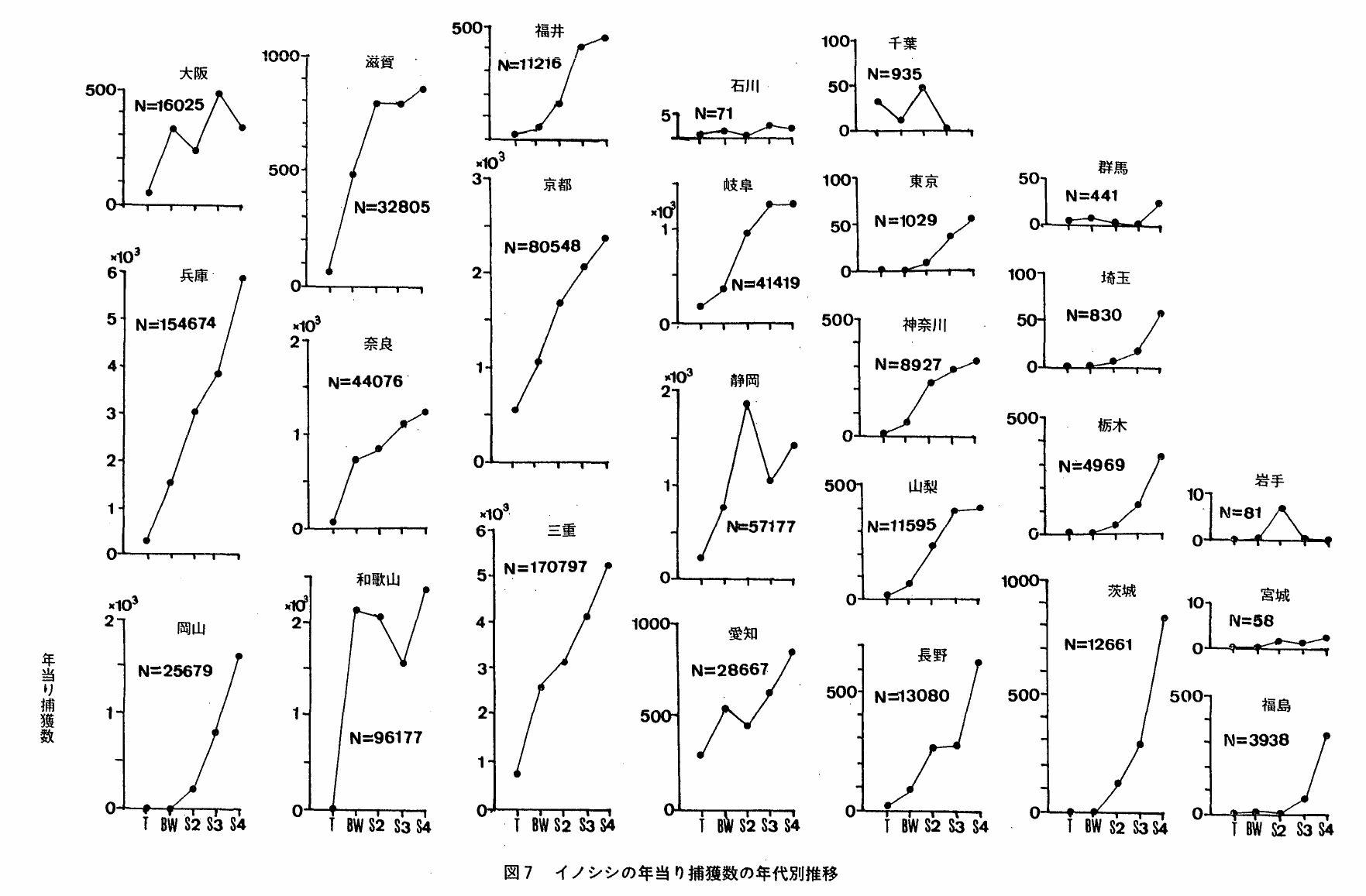
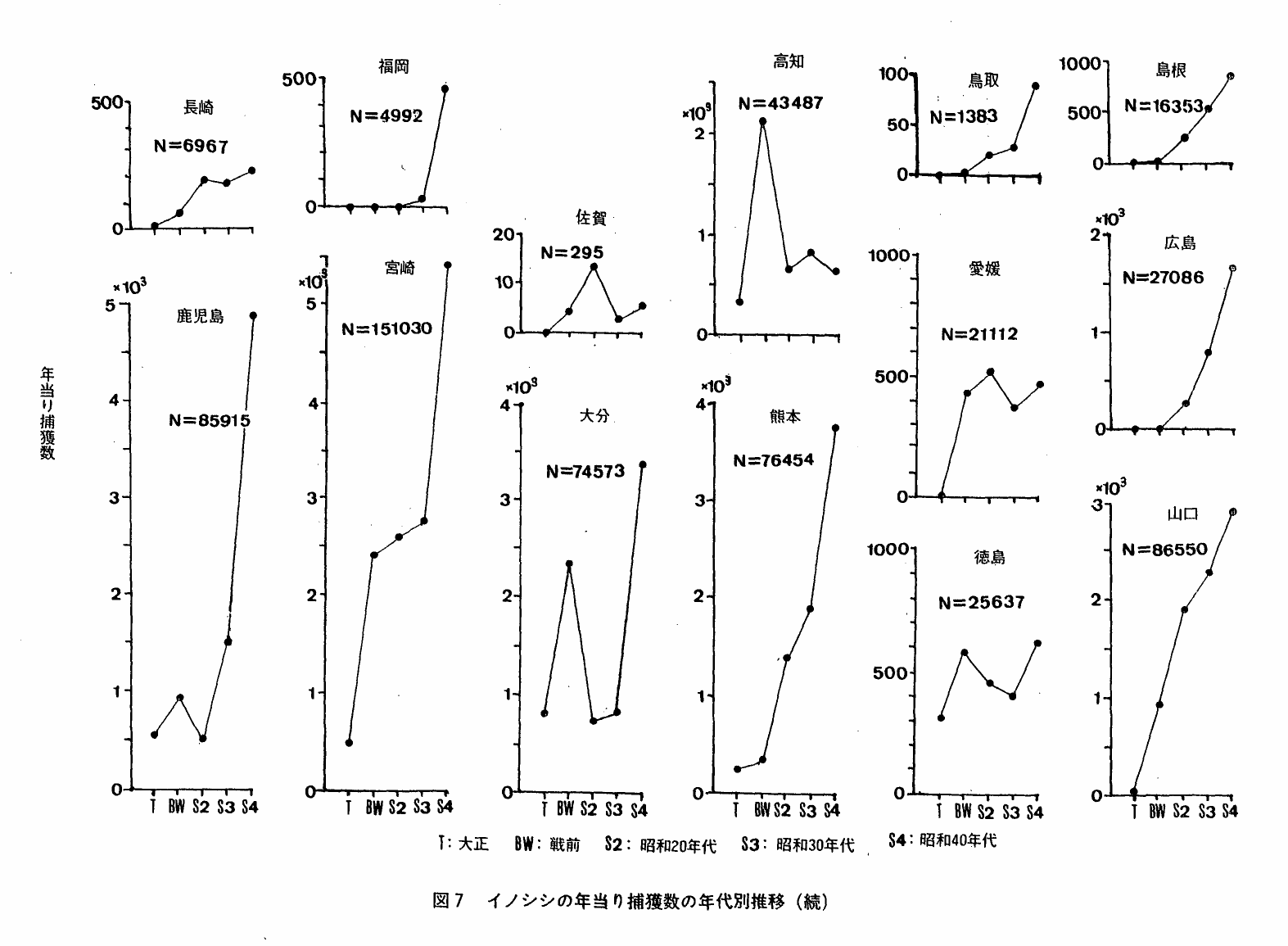
5 本種の捕獲数の推移
捕獲数の変動には,(1)狩猟者数の増加(2)捕獲方法の変化(3)狩猟還境の変化(4)個体数変動,など複数の要因が同時に関係していることが考えられる。したがって,捕獲数の変動だけで本種の動向判定を試みることは出来ないが,他の情報とあわせるならば補助的な情報として使用することが出来ると考えられる。
図7は,還境庁「鳥獣関係統計」にもとずく本種の年当り平均捕獲数の年代区分別推移をみたものである。都府県によって種々なパターンが認められるが,おおよそ次の6種類に区分し得る。
S字型:年当り捕獲数は急増,以後漸増・横這・震動のいずれか。愛媛・福井・長崎・神奈川・和歌山・静岡・岐阜・山梨・滋賀・大阪・奈良
J字型:年当り捕獲数は直接的にまたはJ字状に増加。福岡・鹿児島・熊本・宮崎・広島・岡山・栃木・山口・茨城・福島・東京・埼玉・鳥取・長崎・三重・愛知・兵庫・京都・島根
Eo型:年当り捕獲数は急増,以後減少し振動。高知・佐賀
N字型:年当り捕獲数は急増→減少→急増。大分・徳島
E 型:年当り捕獲数は大きく震動した後ゼロになる。千葉・岩手
lo型:年当り捕獲数は低いレベルで震動。石川・宮城・群馬
6 本種個体群の動向
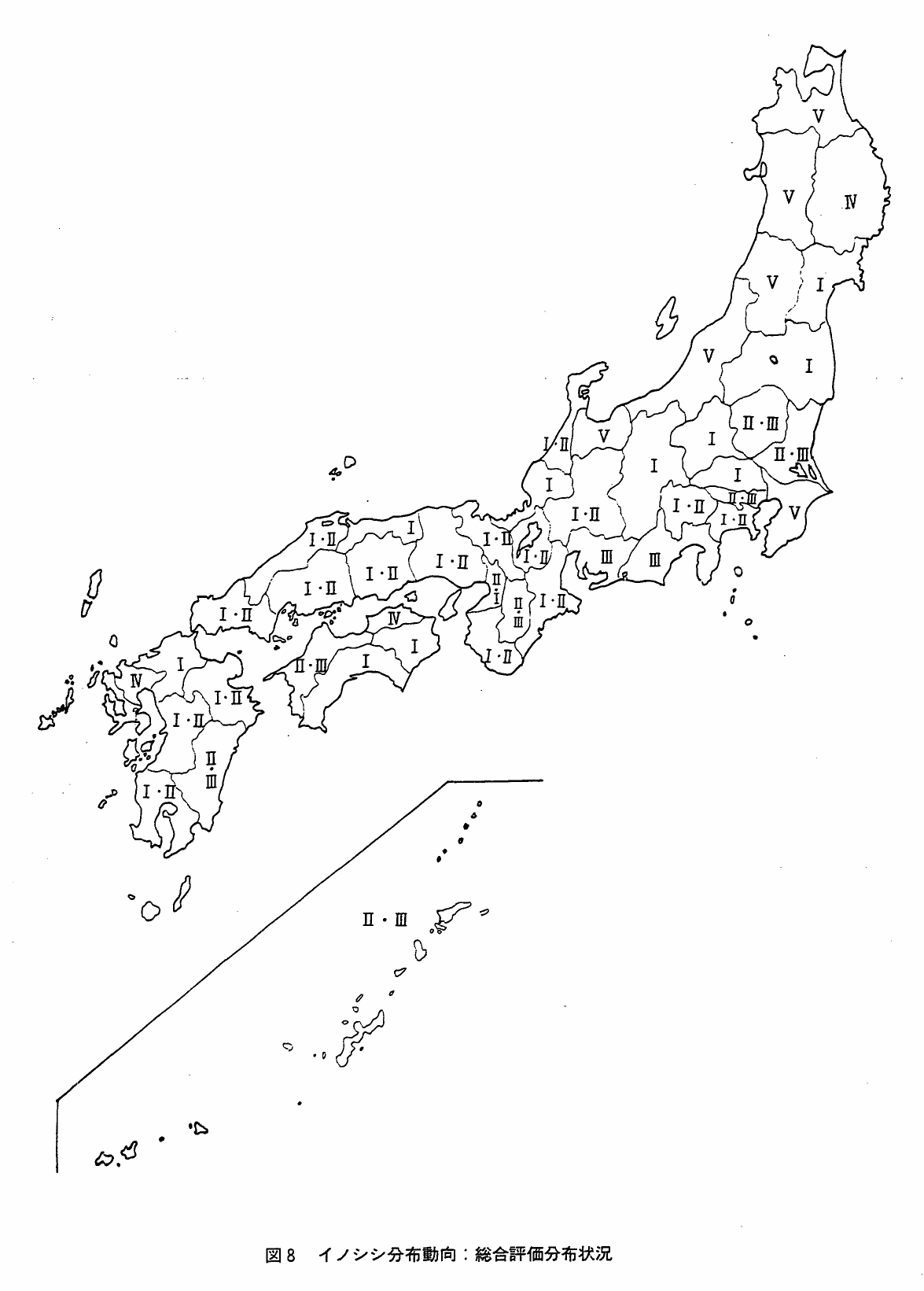
表4は,すでに述べた年当り出現情報数の年代区分別推移パターン,年当り絶滅情報数の年代区分別推移パターン,対森林「9」「6」生息飽和率,年当り捕獲数の年代区分別推移パターン,それに積雪から,イノシシの昭和50年現在の分布動向を総合的に評価したものである。ここでの総合評価区分は,次の6区分とした。
Ⅰ(成長):昭和40年代の出現情報数が絶滅情報数を上まわっていること。出現情報数推移パターンがJ,W,VU,N字型であり,絶滅情報数推移パターンがA,D字型であること。対森林生息飽和率が高くないこと。捕獲数推移パターンがlo,Eo型であること,これがJ,S,N型を示す場合でも昭和40年代の捕獲数が多くないこと。
Ⅱ(横這):昭和40年代の出現および絶滅数に大きな差がないこと。対森林生息飽和率が高いこと。捕獲数がほぼ一定あるいは漸滅傾向が認められること。
Ⅲ(衰退):昭和40年代の出現情報数が絶滅情報数よりもはるかに少ないこと。出現情報数推移パターンがL字型またはA型で,絶滅情報数推移のパターンがJまたはS字型であること。捕獲数推移が上向きであること。
Ⅳ(隣接地域に依存):本種が生息するための還境条件は揃っているが,核となる個体群が存在しない場合。
Ⅴ(分布の可能性小):自然および人為的分布制限要因が強く働いているため,当面分布の可能性はきわめて小さい。
?(不明):評価不能。
上記の区分条件にもとずく評価の結果は表5に示した通りである。但し,ここで行った評価は限られた情報にもとずくものであり,あくまで試行的,目安の域を出るものではない。信頼度を高めるためには,オーソドックスな個体群生態学的手法にもとずく必要があることは当然である。
図8は,上述の本種の分布動向総合評価の分布状況をみたものである。評価Ⅴが東北および北陸地方を中心に分布しているのは当然としても,評価Ⅰがいわゆる分布の周辺部に多いことは注目に値する。
7 本種個体群の地理的分布構造
自然的分布制限要因としての積雪との関係からみるならば,本種の分布中心は無雪あるいは寡雪地帯であり,多雪地帯へむかって分布周辺が拡がっているとみることが出来る。しかし,各種の開発が歴史的に展開されそして集積している今日,自然的分布条件からみた分布中心も,人為的分布制限要因が強く影響しない地域に制限されるとみることができる。そのような地域は,本報ですでに分析したようにまさしく森林区分「9」ないし「6」の地域である。但し,ここでも狩猟圧をはじめとした分布制限要因が働いているから,現在の分布中心はこれらの分布制限要因の影響にもかかわらず強い勢力を有する個体群が生息している地域とみることが出来る。
先に分析した対森林生息飽和率は,このような個体群の勢力を推量するひとつの目安と考えられる。したがって,この生息飽和率が高い地域こそ勢力の強い個体群が分布している可能性が大きい,すなわち分布中心である可能性が大きいとみることが出来る。
すでにみた図2で明らかなように,本州では対森林生息飽和率の高い地域は,近畿地方を中心として中国地方,中部地方へと拡がっているから,これらの地域は分布中心とみなし得る。一方,北陸,関東,東北地方での飽和率はきわめて低く,生息しない地域すら認められるから,これらの地域は分布周辺とみなして差支えない。一方,四国地方は愛媛県での飽和率がきわめて高く,高知・徳島両県がこれに次ぎ,香川県では20%以下ときわめて低くなっている。したがって,四国では,愛媛県を中心とした地理的分布構造を持っているとみられる。九州地方では,宮崎県の飽和率がきわめて高く,隣接する大分,鹿児島,熊本県が次いで高くなっている。そして福岡,佐賀,長崎県の順に低くなっている。当然のことながら,九州地方では宮崎県を分布中心とし,北・西九州を分布周辺とする分布構造となっているとみられる。
図5の出現情報数推移パターンの地理的分布をみると,分布中心とみなされた地域にはW,N,VU型が集中し,それを取り囲むようにJ字型が分布中心から周辺部へと分布している。これは,分布中心の個体群は好適な還境を背景にしているため勢力が強く,多少,分布制限要因が強く働いても,比較的早く回復することを示しているかもしれない。一方,分布周辺のJ字型地域では,分布中心よりもやや遅れて最近になって分布域を拡大しつつあるとみることも出来よう。分布動向の総合評価でもみたように,分布中心では,分布動向総合評価Ⅰ(成長)またはⅡ(横這)であるが,分布周辺ではむしろ評価Ⅰ(成長)が少なからず認められるのは,分布中心と周辺の関係を考える上で重要である。当然分布中心として位置づけられて然るべき高知・静岡両県が総合評価Ⅲ(衰退)であるのは,狩猟圧が強く働いているためではないかと考えられる。これは,狩猟によって絶滅,そして,分布周辺化した千葉県とともに,分布中心が消滅し,分布が切断されていく過程を検討する上で注目に値する。分布周辺にあって総合評価Ⅰ(成長)を示す宮崎・福島・群馬・長野の各県および評価Ⅰ・Ⅱを示す石川・岐阜両県の個体群の動向は,多雪地帯への分布域拡大という点で注目に値しよう。
8 引用文献
朝日 稔,1975,イノシシおよびシカの捕獲と植生区分,生物科学,27(3):159-168。
千葉徳爾,1959,九州北部の野生大型哺乳類分布,地理学評論,32(9):468-480。
千葉徳爾,1964,日本列島における猪・鹿棲息状態とその変動,1964,地理学評論,37(11):575-592。
GILBERT, P. F., O. C. Wallmo and R. B. Gill, 1970, Effect of snow depth on mule deer in Middle Park, Colorado. J. Wildlife Mgt, 34(1):15-22
SOKOLOV, G. A. 1962, (The distribution of the wild boar and the Siberian wild goat in the central part of the Western Sayan and some questions of their ecology, Problems of zoological research in Siberia) from Biol. Abst, abstracted from Referat. Zhur. Biol, 1964, NO. 11236
高橋春成,朝日 稔,1978,イノシシの分布を規制する諸要因について―京都府北部の場合―,「環境科学」研究報告集「日本の歴史的自然としての哺乳類」(四手井綱英編):11-16。
鳥居春己,1978,静岡県におけるサル,クマ,イノシシ,シカ,カモシカの分布と被害状況,研究調査資料,21,静岡県林業試験場。
9 摘 要
環境庁第2回自然還境保全調査の結果にもとづき,イノシシ(Sus scrofa)の地理的分布構造と分布要因を検討した。
本種の多雪地帯への分布境界線は,積雪深30cm以上の1冬あたり日数70日線にほぼ一致した。これを越える多雪地域が多い青森,秋田・山形・新潟・富山県での本種の生息は認められない。本種の生息は森林率40%以上の地域に集中していた。対森林生息飽和率は,近畿・四国・南九州地方で多く,北陸・東北・北および西九州で低かった。これに出現情報数,絶滅情報数を加えて,個体群の分布構造を明らかにし,本州では近畿地方が,四国では愛媛県が,九州では宮崎県が分布中心であること,東北・北陸・北西九州が分布周辺であることを示した。また個体群の動向評価を行い,成長を示す個体群は分布周辺で認められることを示した。
Summary
Factors Affecting the Geographical Distribution
of Japanese Wild Boars
This paper is concerned with the distribution of the Japanese wild boar (Sus scrofa) and the factors affecting this distributin. The data used are from the Second Basic Survey of the Natural Environment. The line indicating 70 days of snow per winter with a snow depth of more than 30 cm coincided almost perfectly with the distribution border of this species. Heavy snow accumulation prevented the boar from inhabiting Aomori, Akita, Yamagata, Niigata, and Toyama Prefectures. This species lives mainly in regions of more than 40 percent forestation. The frequency of habitation of this species in its favorite forests was high in the Kinki, Shikoku, and southern Kyushu Regions and low in Hokuriku, Tohoku, and northwestern Kyushu. Analyzing imformation on the appearence and extinction of boars, the geographical centers of distribution are considered to be located in the Kinki district in Honshu, in Ehime Prefecture in Shikoku, and in Miyazaki Prefecture in Kyushu. Tohoku, Hokuriku, and northwestern Kyushu were considered to be peripherral areas where local populations may possibly have developed. (Kunihiko TOKIDA, and Naoki MARUYAMA)
付表2 イノシシの生息・絶滅区画数およびそれらの全区画数に対する割合
目次へ
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}