3 ツキノワグマの分布について
花 井 正 光
1 はじめに
全国を対象としたツキノワグマの分布現況についての最初の調査は,アンケートの郵送による方法をとって実施された(哺乳類分布調査科研グループ,1979)。この際得られた分布図の精度を高めるためには,補完的調査としての聞きとり調査を必要とすることが,部分的な両調査の実施により指摘されていた。第2回自然環境保全調査の対象となった哺乳類の分布調査が聞きとりアンケートを調査方法としたのは,上の経緯からして必然的であったし,44,853人に及ぶ回答者から情報を得た結果,前者の5kmメッシュのグリッド・システムによる分布図を補完して余りあるものとなった(環境庁,1979)。
ここでは,ツキノワグマの分布域を地方別にその広がりの現況と変動をながめ、分布成因としてのブナ,ミズナラを指標種とする温帯落葉樹林の分布との関連性についてもふれることにしたい。また,分布域における地域的または局地的なツキノワグマの分布についても,生活場所(ハビタット)の選択を土地利用として生態的な分布構造の側面から考察を加えたので報告し,今後のツキノワグマの研究調査の展開素材としたい。
報告に先立ち,秋田県下で害獣駆除によるツキノワグマ捕獲地点をまとめた未発表の分布図の引用を認められた秋田県林政課に対し感謝の意を表しておきたい。
2 方 法
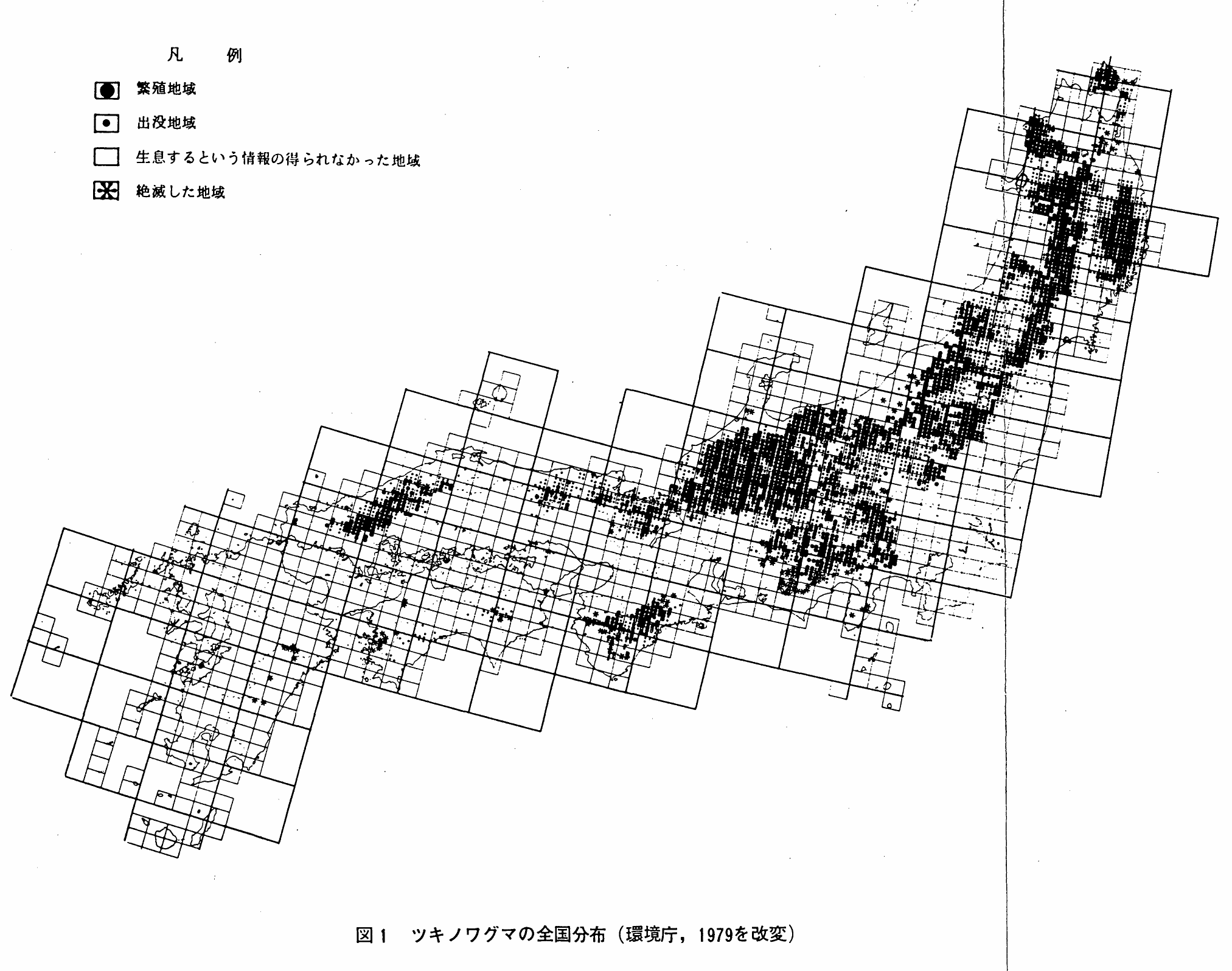
1kmメッシュによる生息区画数,絶滅区画数,季節別出現区画数など都府県ごとの電算機集計値(付表)とそれぞれの区画を2kmメッシュで電算機によりえがいた分布図のほか,既存資料として5kmメッシュによる分布図(図1)や生息および絶滅区画数を使用した。
また,ツキノワグマの分布と植生との関連性を示すために,1kmメッシュにより都府県別に集計され植生区分を利用した。
このほか環境庁によって毎年発行される鳥獣関係統計から都府県ごとの本種の捕獲数を抜すいし,その推移を参考資料とした。
3 結果と考察
(1)分布現況
ツキノワグマの分布の現状を概略して述べれば,中部地方以東の山岳地帯に連続分布域がみられ,以西では対照的に不連続分布となっている(哺乳類分布調査科研グループ,前掲;環境庁,前掲)。
ここでは,今回あらたに作成された1kmメッシュによるデータを使用して,上の分布現況について地方ごとに細分してながめる。
東北地方:この地方は,中央部の脊稜山地を中心に隣接する主な山地のほとんどの地域にわたって連続して分布域が広がっている点で,他の地方にはみられない特徴を有している。即ち,奥羽山脈を軸に白神山地,北上山地,丁岳山地,出羽山地,朝日・飯豊山地および帝釈山地などに連続分布がみられる。
このほか下北半島にも分布するが,この分布域は,半島の基部が100m以下の低地となっており,これにより隔離されているとみることができるが,一時的に出没する区画が少数ながら点在することもあって,隔離がどの程度成立しているかは判断できない。また,北上山地の分布域も北上川の流路にそって奥羽山脈の連続分布域と隔離されている可能性を分布図(環境庁,前掲)は示しているが,北部地域で一部連続する区画がみられることから,この隔離は強くはないかもしれない。
以上のように一部でやや不連続な分布域が存在するものの,東北地方におけるツキノワグマの分布域はほぼ連続しているとして大過なかろう。
東北6県における生息区画率は,5kmメッシュでは総区画数2,746.5に対し1,442で52.5%,約半数の区画で生息情報が得られている。1kmメッシュでは総区画数66,216となり,このうちの7.9%が生息区画となった。両メッシュで生息区画率に大きな差を生じているが,これは聞きとり調査を5kmメッシュを用いて実施した結果を,1kmメッシュに置換したことによりもたらされたもので,他の地方についても同じく差は大きいものとなった。いずれにしても,地方別にみた場合,生息区画率のもっとも高い地方となっている。
関東地方:東北地方に続く中央脊稜山地およびその隣接山地である三国山地,越後山地,関東山地に連続分布域がみられる。隔離された分布域はみられないものの,茨城,千葉両県のごとく全く生息区画がないか,生息区画率が低い都県を含む結果,全体としての分布域の広がりはさほど大きくはない。
5kmメッシュによる生息区率は,総区画数1,247.5に対し297で23.8%であり,1kmメッシュでは,総区画数30,284に対し,1,092で3.6%となった。
中部地方:この地方の分布域は前の東北および関東のそれに隣近かつ連続している。南北両アルプス,中央アルプスはじめ,飛騨山地,両白山地などが主な分布域となっている。
この地方には長野,岐阜,福井各県のように,5kmメッシュでは70%を越える生息区画率を示し,その結果東北地方にほぼ近い連続した分布域の広がりをもっている。ことに長野,岐阜両県下での生息区画は多く,東北地方の岩手,山形,秋田などとほぼ同様で,ツキノワグマ分布域中,もっとも高い生息区画率を示すグループを形成している。
中部地方全体では,5kmメッシュの場合総区画数2,770.5に対し生息区画数は1,323,1kmメッシュの場合は63,391と4,116で,生息区画率はそれぞれ47.8%と6.5%であった。
近畿地方:両白山地に隣接して比良山地や丹波山地など日本海側に面した山地に連続する分布域があるが,中部地方以東に較べてその広がりは著しく狭いものとなっている。しかも,この分布域をもって東北地方からの連続分布の西限を画している。また,このほか紀伊山地に,他のどの分布域とも繋がらない隔離分布域が存在するのも中部地方以東ではみられないことで,この地方でのツキノワグマの分布を特徴づけている。
生息区画率は,5kmメッシュの場合総区画数1,367.5に対し270.5で19.8%,1kmメッシュでは30,354と972で3.2%となり,関東地方に次いで低い値となっている。
中国地方:近畿地方西部の分布域が中国山地の東部まで連続しているが,岡山,鳥取および兵庫の県境付近までで,それより西には広がっていない。したがって,この地方の実質的な分布域は,中国山地西部に位置する冠山山地周辺にみられるだけであり,紀伊山地同様,隔離された孤立分布域となっている。
生息区画率は,5kmメッシュの場合総区画数1,420に対し227で16%,1kmメッシュでは29,657に対し779で2.6%となっている。この値は近畿地方よりも一層小さなものとなっている。
四国地方:南北に二分して東西に走る四国山地の東部と西部にわずかずつ生息区画が点在するのみで,これまでみてきたいずれの地方にもなかったきわめて貧弱な分布が示されている。
他の地方で分布域のほぼ全域にわたって得られている繁殖に関する情報が皆無であり,季節的または一時的な出現の情報しかなかったことも,上の貧弱な分布現況と無縁ではなく,地域個体群の消滅が懸念される。
5kmメッシュにおける生息区画はわずかに26で,総区画数の3.9%に過ぎず,1kmメッシュでは0.3%とさらに僅少なものとなる。
九州地方:この地方では分布域は現存せず,すでに絶滅したものとみてよい。1kmメッシュでは宮崎県下に4区画のみ生息区画がみられたが,いずれも生息に関する情報が年代などを欠いており,実質的には生息区画とみなさずともよいものである。
(2)分布域の変動
ツキノワグマの分布現況は上にみたとおりであるが,このうち分布域の拡大が近時になって引きおこされたはっきりとした例はいずれの地域においてもみられないようである。逆に,地域的な絶滅により分布域が消滅したり,地域個体群の絶滅の可能性が高い分布域の存在や,不連続な隔離分布域が生じるといった退行的な分布域の変動が明らかになった。
このほか,今回の調査では絶滅に関する情報が得られていないが,自然環境からかつては分布域であったと考えられる地域が幾つか存在することも示唆された。
絶滅による分布域の消滅:九州地方があげられることは既に述べた。今回の調査では,5kmメッシュで大分県は3区画に8件,宮崎県は4.5区画に36件それぞれ絶滅情報が得られた。これらは九州山地の祖母・傾山系と宮崎県中部の山地に比較的限定されている。また,絶滅の年代としては,明治,大正とする情報が宮崎県で66%,大分県で50%あった。
狩猟統計によれば,昭和26年に大分県で3頭の捕獲が記録されて以後捕獲は途絶えている。また,大正12年以降大分県ではわずかに年1~4頭の記録が前後7年とびとびにみられるのみで,宮崎県を含めて他県にはみるべき記録はない。このように九州地方ではツキノワグマの捕獲数はごく僅少であった。
小野(1977)は,祖母・傾山系における本種個体群の崩壊が昭和10年を境におこったと推察し,崩壊の前後の個体群サイズとして100頭内外および10~20頭程度を仮定している。また,千葉(1975)はこの地方で,クマを捕獲すると崇りがあるとして,1頭ごとに暮石を建る狩猟習俗を採録している。
九州山地では,後述するように生息場所の広がりが他地域に比較して著しく狭かったとすれば,地域個体群の規模も小さなものであり,いわば遺存的な性格をもった分布域として存在していたと言えよう。そして,実質的に地域個体群が絶滅した時期は昭和以前を想定して大過ないものと思われる。
絶滅に瀕している分布域:このケースとしては四国地方があげられる。四国山地には東西に二分して生息域がみられるものの,ともに生息区画は僅少で,しかも同時に絶滅情報をも含んだ生息区画が多く,繁殖を裏付ける情報が皆無であることなどから,個体群としての存続が危ぶまれる状況にある。
徳島県の剣山地と高知,愛媛県境の両生息域を比較すると絶滅情報を含んだ生息区画が後者で多い。また,後者では生息区画がまばらに散在することから,地域個体群としては後者の方がより小さいものと推察できる。このことは両地区を含む徳島県と高知県における近年の捕獲数にも示されている。
狩猟統計によれば,昭和53年までの10年間の年平均捕獲数は,徳島4.8,高知1.6である。
九州地方と同じく,生活場所の広がりが狭く,遺存的な分布域の性格をもつことから,絶滅に繋がりやすい状況にあると考えられる。
なお,四国地方のツキワグマの分布を述べたものに古屋(1978)があるが,今回の結果と大きく違ってはいない。
隔離分布域の出現:紀伊山地と中国山地西部にほぼ同規模の分布域が存在する。
明治時代以降の絶滅情報を聴取した今回の調査で,岡山,鳥取県境を主体とした中国山地の中部と紀伊山地に繋がる鈴鹿,布引および高見の各山地には絶滅情報がない。したがって,両隔離分布域をもたらしたこれらの地域の分布域の消滅,即ち切れた分布域となったのは明治以前のことと推察される。
両隔離分布域が出現するに至る過程については,切れた分布域における生活場所としての潜存性,自然環境の人為改変,狩猟圧などから検討しなければならないが,いずれも今後に残される課題である。ただ,中国山地については地理学の立場から,早い時代からの強い人為のおよび方により自然環境の改変が著しかった(例えば,千葉,1973;安田,1980など)という,ツキワグマの地域的絶滅との関連性において興味ある指摘がなされている。
なお,両隔離分布域では,いずれも近年の奥地自然林の人を造林化(拡大造林)による分布域の縮小と分断の進行およびそれに伴う地域個体群の弱小化が指摘されている(高橋,1979;和歌山県,1979)。
かなり古い時代の消滅分布域:九州山地の分布域が事実上昭和の初期に消滅したのに対し,少なくとも明治以前に地域的に消滅したと考えられる分布域がある。
今回の調査では,絶滅情報が得られなかったものの,東北地方や近畿および中国地方で連続分布域に接し,主として植生からみる自然環境も潜在的に生息が可能であると考えられる地域では,かつてはツキノワグマが分布していたに違いない。
紀伊半島および中国山地の切れた分布域の存在については既に述べたが,このほか東北地方で津軽半島と阿武隈山地がこの例として挙げられる。いずれも絶滅の原因と時代は不明であるが,この問題を解明するには,ツキノワグマの生活史はもちろん,例えば餌の量や質の年変動と関連した個体群動態や狩猟在との関係などが検討されねばならない。
以上は比較的広域な分布域,即ち地理的な分布域についてとの変動をみたのであるが,分布域を地域個体群を単位にした局地的または地域的な分布をみた場合,分布の変動が生じているケースも多い。
主として天然林の伐採とその後の針葉樹の人工植栽によるツキノワグマの生活場所の環境改変を直接的原因として,周辺地域への移動,分散が生じ,農林産物への加害の増大を招く結果,害獣として駆除される。この一連の過程による地域個体群の崩壊が指摘されている(古林ほか,1974;同,1978;野崎ほか,1979など)。狩猟統計によれば,昭和40年前後を境にして害獣駆除による捕獲数が猟期中のそれを上まわるに至っていることも上の傾向を明示するもので,今後さらに分布域の各地で局地的絶滅の進行が懸念される。
(3)分布成因としてのブナ帯自然林
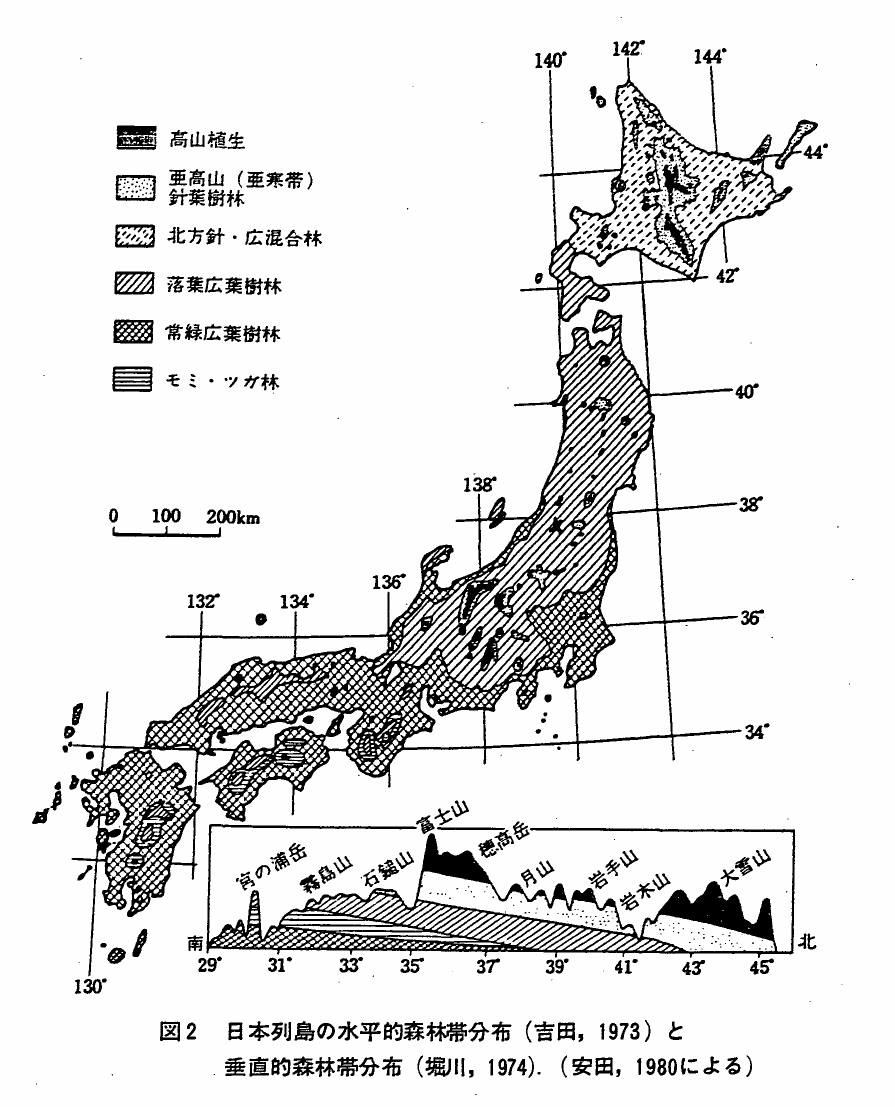
これまでみてきた地方別のツキノワグマの分布は,一見して,本州,四国および九州における落葉広葉樹林の水平的分布とよく一致したものになっている。安田(前掲)より引用した図2にみる水平的森林帯の分布図と5kmメッシュによるツキノワグマの分布(図1)を重ねてみればその整合性は明らかである。
また,落葉広葉樹林に接したコナラ,クリ,シデ類,モミ,ツガなどからなる森林を吉良(1971)は暖帯落葉樹林として区分したが,本森林帯が中部地方から東北地方の内陸部に現存することをあわせると,これら両森林帯でツキノワグマの分布は一層よくカバーすることができる。なお図2には緯度にそった垂直的森林帯が一緒にされているが,先にみた西日本でツキノワグマの分布域が狭少になることは,生活場所としての落葉広葉樹林帯の狭少さと係っていることをよく示すものである。
図2 日本列島の水平的森林帯分布(吉田,1973)と垂直的森林帯分布(堀川,1974).(安田,1980による)
表1 1kmメッシュによる地方別区画総数およびブナ帯区画数とツキノワグマの生息区画数
本種の生活場所がブナ,ミズナラが優占する落葉広葉樹林に求められていることの指摘は少なくない(例えば,鳥居,1974;渡辺,1974;千葉,1975;花井,1977;NOZAKI et.al.,1980など)。また本種の亜種であるソ連極東南部に分布するウスリークロクマやインド,カシミール地方に分布するヒマラヤグマも,ツキノワグマと同じく落葉広葉樹林を生活場所としていることが報告されている(BLOMLEI,1965;SCHALLER,1969)。
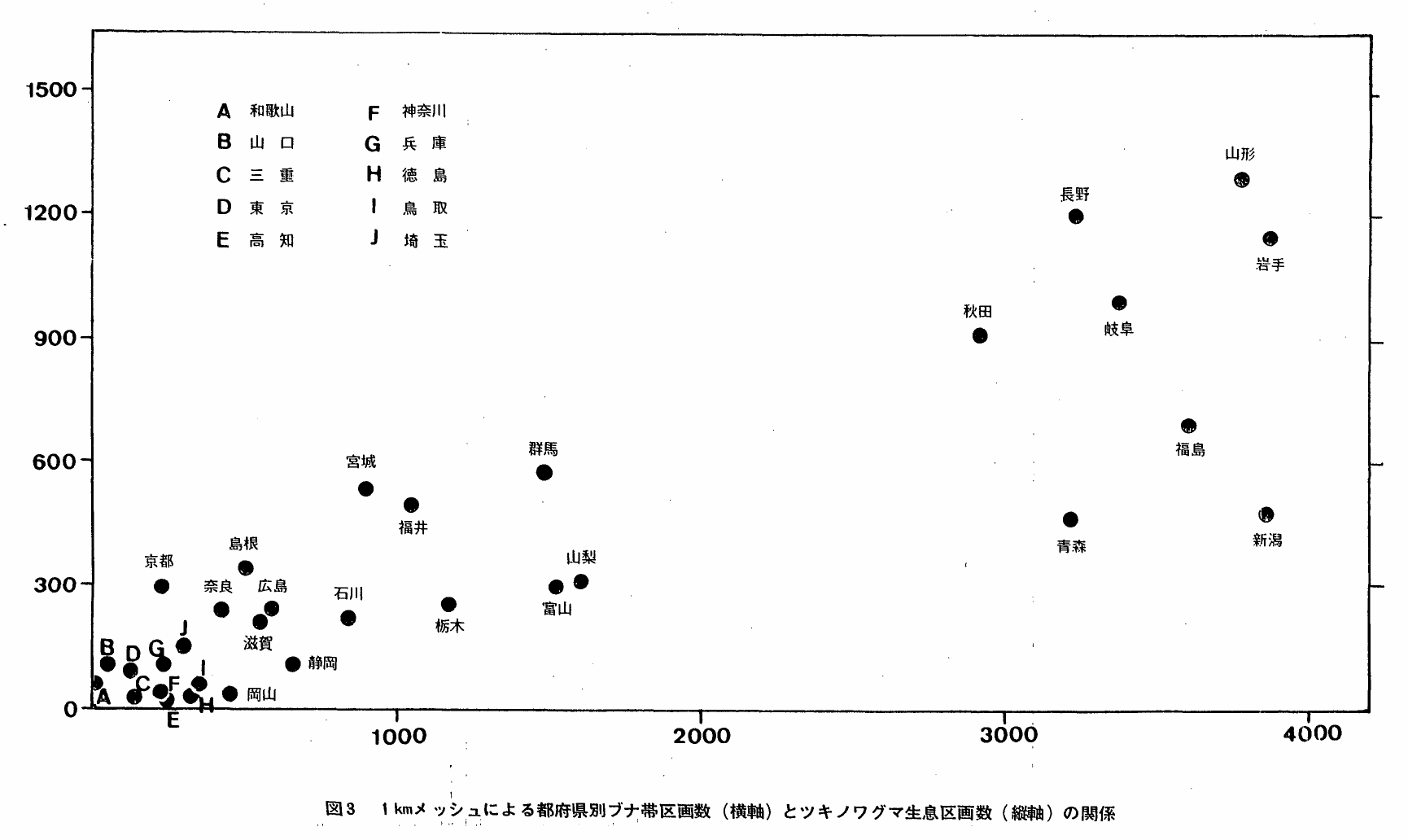
ところで,地方別および都府県別にブナ,ミズナラの森林帯面積とクマの分布面積の関係をみたのが表1と図3である。森林帯面積は第1回自然環境保全調査の成果(環境庁,1976)により,ミズナラ―ブナクラス域の自然および代償植生に区分された1kmメッシュの区画数を全ての群落について総計することで求めた。
地方別にみると1kmメッシュ総区画数に占めるブナ帯区画数およびツキノワグマ生息区画数の割合は同一傾向を示し,ブナ帯区画率の順と生息区画率のそれとは同一となっている。このことは両者の間に正の相関関係が成立することを示唆するものである。そこで,都府県ごとにブナ帯区画数に対する生息区画数をプロットしてみると,おおよその傾向として上の関係が示された,これにより,概して東日本で濃く西日本で薄いツキノワグマの分布のパターンが植生から説明することができる。
しかしながら,分布域の広がりと生活場所としてのブナ帯の広がりの間におおまかな正の相関関係が見い出されるとはいえ,都府県別にはもちろん,表1にも示されるとおり隔離分布域をもつブナ帯域の狭い近畿や中国地方が,広い連続分布域をもつ東北や中部地方よりも逆に高い率で分布しており,両者の関係が単純に成立しているものではないことを同時に示している。本種を含む大型哺乳類の全てが時,所を問わず狩猟対象であったし,自然の人為による変改は所によってさまざまであるから,地域によって現在のツキノワグマの分布が一様でないことは当然のことかもしれない。分布はすぐれて歴史の所産であると考えられる。このことはしかし,上でみたツキノワグマの分布域とブナ帯の広がりの間には,本種が生活場所をブナ帯に求めるゆえにパラレルな傾向が成立することを否定するものではない。
なお,北海道の南部,渡島半島にもブナ帯が現存するが,ツキノワグマの分布をみないのは,地史的な背景が原因となっていると考えられている(新妻,1972)。
(4)局部的な分布構造
ツキノワグマの分布域がブナ帯自然林のそれと重複していることをみた。この分布は水平的ないしは地理的な分布圏ともいえる広域的な側面を有するが,分布域には別の側面も包含されている。それは,分布域はいくつもの地域的ないしは局地的な分布範囲をもって構成されているということである。この分布範囲は,一般的に生活場所の非連続性に対応して形成されると考えられる。ここでは,これらの分布範囲におけるツキノワグマの分布構造について述べるが,それは各々の分布範囲は地域個体群を単位として含んでおり,この個体群の土地利用を生態的側面としてみることでもある。
本調査では,目視の季節をも同時に聞きとることとされたが,その結果本種については秋における目視情報数が,生息情報の著しく少ない一部の県を除いて,ほとんどの都府県で夏,秋よりも圧倒的に多かった。例えば,青森県では生息情報数640のうち秋の目視によるものが60%に及ぶのに対し,夏と冬はそれぞれ15%と8%であった。1kmメッシュの結果で言えば,生息区画数512のうち65%におよぶ区画で秋の目視情報があり,夏および冬は18%と10%であった。秋に目視が多くなるのは,クマがこの季節に人里に近づくことによるもので,この傾向は本種の分布域のほぼ全域にみられ。
図3 1kmメッシュによる都府県別ブナ帯区画数(横軸)とツキノワグマ生息区画数(縦軸)の関係
一方,農作物への食害を理由に害獣駆除の対象として夏から秋に捕獲されることが多いことは前にも述べたとおりであるが,こうした人里またはその隣接地域での捕獲も上の現象の結果であろう。
以上のことから季節的または一時的にクマが通年の生活場所から移動する現象は一般的で,移動がみられる地域を分布周辺地域として既年生活する分布中心地域と区分することができよう。ここで季節的および一時的な移動の区別は,前者が恒常的な現象であるのに対し,後者は変則的で土地利用形態から言えば結びつきのより弱い空間ということができよう。東(1978)は,分布中心地域および分布周辺地域に対し定住域および一時的分散地域として呼びわけている。
亜高山帯および高山帯が存在する地域では,季節的な移動が高海抜地でも観察されている(羽田ほか,1959;鳥居,1975;花井,1978a)。したがって,地域的にツキノワグマの分布をみるときには垂直的な構造を認めることができる。
また,季節的な移動は餌をめぐる土地利用として生活様式に組み込まれているものと考えられるが,繁殖をともなう分布中心地域では密度調節機構としての機能も予想され,分布周辺地域が安定的な地域個体群の維持に重要な役割を果していると思われる。
表2 中部地方におけるツキノワグマの生態的分布(土地利用)の模式的区分
以上の考え方にもとづき,ツキノワグマの地域個体群の分布構造を模式的に区分してみたのが表2である。中部地方を対象として垂直的森林帯により環境を区分し,それぞれの区分帯に対し採食,繁殖,越冬などのための土地利用が時間的にどの程度依存性をもっているかによって生態的分布の区分を試みたものである。
ツキノワグマの地域的な生態的分布もしくは土地利用は,社会構造や個体群の動態との関係において明らかにされねばならないが,そのためには移動や分散様式を生活環境の季節的および年次的変動との関係に注目した野外調査の必要性が指摘されながら,この面での研究は従来ほとんど取り組れておらず今後の課題として残されているのが,現状である。
(5)分布調査から保護管理策の具体化へ
聞きとりによる今回の調査では,先に哺乳類分布調査グループ(前掲)の実施した郵送アンケートによる結果とあわせて,全国規模での大型哺乳類の分布現況の把握に成功したと言えよう。
既に絶滅した分布域がある一方,絶滅に瀕している分布域や絶滅を引きおこしやすい隔離分布域の存在などが明示されたわけであるが,これらはツキノワグマの保護管理策の早急な確立と実施の必要性をも明白にしたと考えるべきであろう。
本種の地域個体群の動態を把握することは方法上容易でないが,個々の分布域における本種の適切な保護管理を志向するには是非とも解決しなければならない問題である。調査方法の開発を含めて調査研究が展開されることになるなら,今回の基礎調査は本種の生態解明のみならず保護管理策の確立へ導くものとしてその意義が高く評価されることになろう。
地域個体群を単位とした動態研究は初原的ではあるが試みられてはいる。捕獲個体を使用した年齢査定により個体群の年齢構成を把握したり,捕獲努力量と捕獲数との関係を経年的に調査することである程度の動向を判定することができる(花井,1978b;HANAI,1980)。この場合,捕獲個体は全数を供試材料にすることが肝要であるが,現状では制度上の保障もなく材料の採集は困難である。しかしながら,上述の早急な保護管理策を要する分布域については,年齢査定用の頭骨もしくは特定の歯牙や捕獲データなどの採集システムの制度的な整備の検討が急がれねばならない。
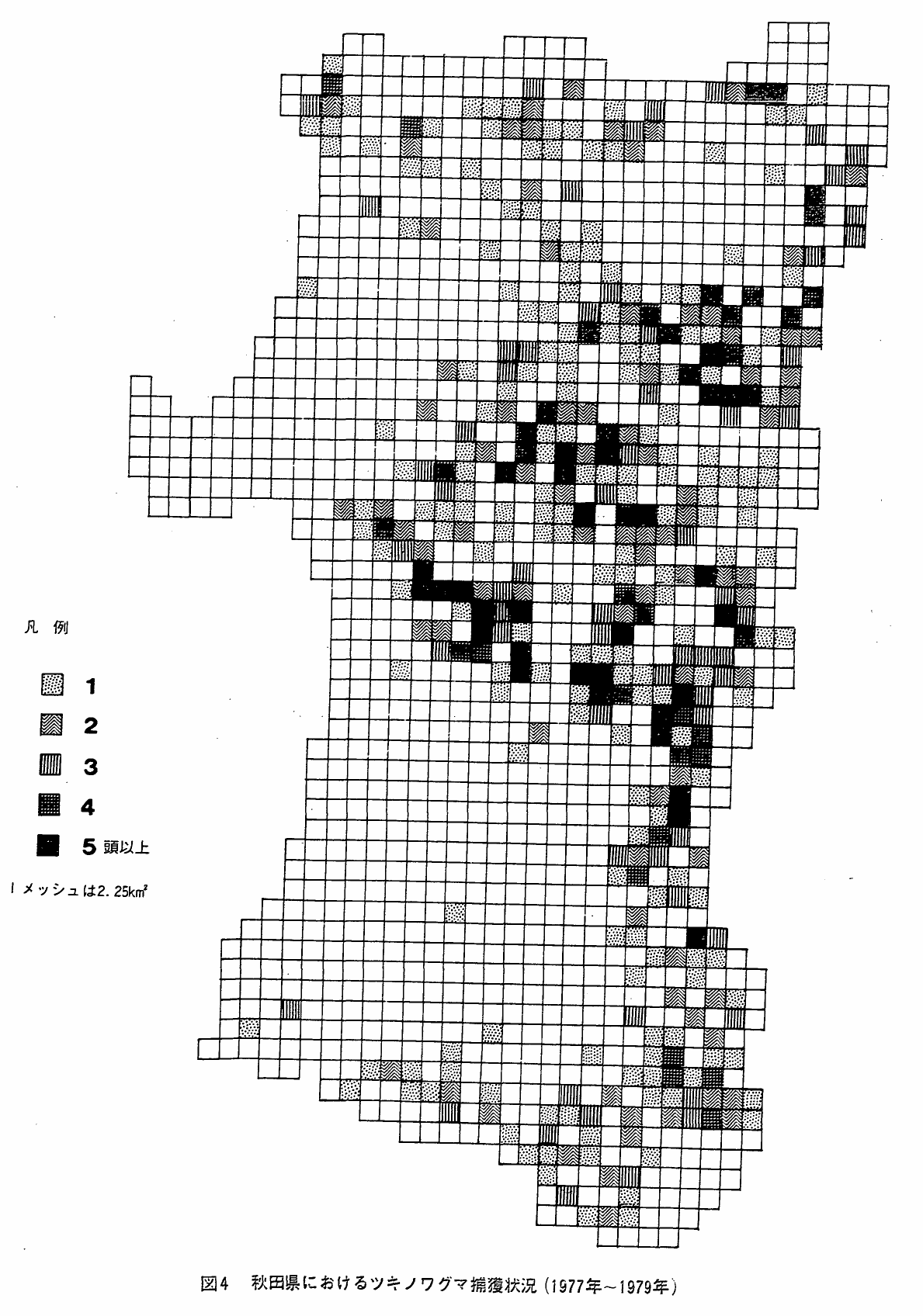
ところで,図4は秋田県林政課が独自に設定したグリッド・システム(6kmメッシュ)を使って,昭和52年から54年の間の害獣駆除によるツキノワグマ捕獲地点をメッシュごとに累積して示したものである(未発表)。秋田県について今回の調査で得た分布図とこの分布図はよく一致しているのである。このことは,狩猟および有害鳥獣駆除に関する従来の行政システムによっても,分布の変動はもちろん個体群動態についての一次資料を集積できることを示すものである。この際,末端の機関としては,県レベルではなく,実質的な鳥獣行政の未端機関となっている県の出先機関が有効かつ実際的であると考えられる。
こうして分布図を経年的に作成することができれば,今回重ねあわせを試みた植生との対比により,ツキノワグマの分布変動を追跡でき,自然環境保全基礎調査の機能的な活用が期待できよう。
図4 秋田県におけるツキノワグマ捕獲状況(1977年~1979年)
4 引用文献
東 滋 (1978) 奥美濃のクマとクマ猟。「環境科学特別研究 日本の歴史的自然環境としての哺乳類 研究報告集」(四手井綱英編),58-66。
BLOMLEI,G.F. (1965) 南部シベリアのヒグマとツキノワグマ。〔藤巻裕蔵・新妻昭夫訳(1972)北苑社〕。
千葉徳爾 (1973) はげ山の文化。233PP.,学生社,東京。
―― (1975a) 狩猟伝承。325PP.,法政大学出版局。
―― (1975b) 蔵王山東麓における野生大形哺類の分布およびその変動について。東北地理,27(2):74-81。
古林賢恒・丸山直樹・三浦慎悟 (1974) 南アルプス,大鹿村における自然破壊――森林の獣害から。ミチューリン生物学研究,11(1):2-5。
古林賢恒・宮本雅美・水口民夫・渡辺弘之 (1978) 京都府での捕獲と分布域からみたツキノワグマの拡散的絶滅過程。「環境科学特別研究 日本の歴史的自然環境としての哺乳類 研究報告集」(四手井綱英編),41-48
古屋義男 (1978) 四国における中大型哺乳類5種の分布――アンケート調査による――。高知女子大紀要 自然科学編,26:13-19。
花井正光 (1977) 石川県の哺乳類。「石川県の自然環境 第3分冊 鳥獣」,149-198,石川県。
―― (1978a) 白山高山帯の哺乳類相。石川県白山自然保護センター研究報告,4:83-92。
―― (1978b) ニホンツキノワグマとリュウキュウイノシシの齢組成にみられる年次変動について。個体群生態学会会報,31:16-18。
HANAI,M.(1980)Population characteristics of the Japanese black bear in Hakusan National Park,Japan.「Bears――Their Biology and Management,Bear Biology Association Conference Series No.3」(Edited by Martinka,C.J.and K.L.McArthur),63-66.
羽田健三・千葉淋司 (1959) 針ノ木岳における大型哺乳類の社会生態学的研究。「針ノ木岳」(大町山岳博物館編),63-72。
哺乳類分布調査科研グループ (1979) カモシカ・シカ・ヒグマ・ツキノワグマ・ニホンザル・イノシシの全国的生息分布ならびに被害分布。生物科学,31(2):96-112。
環境庁 (1976) 自然環境保全調査報告書。399pp,.。
―― (1979) 動物分布調査報告書(哺乳類)全国版。91pp.,。
吉良竜夫 (1971) 生態学からみた自然。295PP.,河出書房新社。
新妻昭夫 (1972) 北限のツキノワグマとツキノワグマの北限,哺乳類科学,25:1-12。
野崎英吉・古林賢恒・丸山直樹・常田邦彦・遠竹行俊 (1979) 関東地方におけるツキノワグマの分布――アンケート・聞きとり調査による――。哺乳動物学雑誌,8(1):1432。
NOZAKI,E.,S.AZUMA,T.AOI,H.TORII,K.MAEDA and T.ITO(1980)Food habits and habitat utilization of Japanese black bears,(manuscript for proceedings of the Fifth International Conference on Bear
Research and Managenient)
小野勇一 (1977) 九州のニホンツキノワグマ。Vulpes,5.
SCHALLER,G.B.(1969)Food habits of the Himalayan Black Bear in the Dachigam Sanctuary,Kashimir.J.Bombay Nat.Hist.Soc.,66(1):156-159。
高橋春成 (1979) 西中国山地におけるツキノワグマの分布動向と植生変化。地理学評論,52(11):635-642。
鳥居春己 (1074) 静岡県北部のツキノワグマ。哺乳動物学雑誌,6(2):94-99。
―― (1975) 大井川流域のツキノワグマ。「南アルプス・奥大井地域学術調査報告書」,56-60。
和歌山県 (1978) 動物分布調査報告書(哺乳類)。
渡辺弘之 (1974) ツキノワグマの話。172PP.,日本放送出版協会。
安田喜憲 (1980) 環境考古学事始。270PP.,日本放送出版協会。
吉岡邦二 (1973) 植物地理学,生態学講座12。84PP.,共立出版。
5 摘 要
第2回自然環境保全調査(1978)の一環として実施された哺乳類分布調査の結果にもとづき,ツキノワグマの分布現況,分布の変動,分布の成因および分布構造について考察した。
中部地方以東で連続分布するのに対し,以西では隔離された狭い分布域となり,ことに四国地方ではきわめて狭少化したものとなっている。
九州地方の分布域はすでに消滅しており,きわめて狭あい化した分布域しかみられない四国地方では消滅の可能性が高い。
隔離分布域を生じている地方や連続分布域の一部で明治時代以前のかなり早い時代の地域的な分布域の消滅が示唆された。
ツキノワグマの分布域とブナ,ミズナラ類を主体とする落葉広葉樹林の分布がよく一致すること,両分布の面積の間におおまかな正の相関関係が認められることから,ツキノワグマの分布はこの森林帯の分布に規定されていると考えられた。
ツキノワグマの分布域を地域的または局地的にみると,森林帯が垂直的な分布構造をもつことに関連して,土地利用形態に応じて分布の中心地域と周辺地域が形成されると考えられた。
絶滅に瀕している四国地方や中国,近畿地方の隔離分布域に対しては,地域個体群の動態指標を得るために捕獲個体を使用する制度の検討を含め,早急な保護管理対策の着手が望まれる。
Summary
Status of distribution of the japanese black bear (Selenarctos thibetanus japonicus) was surveyed by grid system. In the eastern part of Honshu distribution area spreads out continuously. By contrast, there are only two isolated distribution areas in western part. It is diminishing in Shikoku and has already vanished in Kyushu.
Bear distribution and Querco-Fagetea region frequently coincide. A correlation was recognized between the two proving that natural deciduous forest of Querco-Fagetea region is prefered as the habitat.
Seasonal habitat utilization by local population forms the central area and peripheral area.
付表2 ツキノワグマの生息・絶滅区画数およびそれらの全区画数に対する割合
目次へ
{kind=link}
{kind=link}
{kind=link}
{kind=link}