�@
�������z�����̈�Ƃ��čs��ꂽ���E��^�M���ނ̕��z�����őΏۂƂȂ����M���ނ́A�j�z���U���iMacaca�@fuscata�j���i�P�j�A�V�J�iCervus�@nippon�j���i�Q�j�A�c�L�m���O�}�iSelenarctos�@thibetanus�j�A�q�O�}�iUrsus�@arctos�j�A�C�m�V�V�iSus�@scrofa�j���i�R�j�A�L�c�l�iVulpes�@vulpes�j�A�^�k�L�iNyctereutes�@procyonoides�j����уA�i�O�}�iMeles�@meles�j�̂W��ł���B
�����̚M���ނ́A�����n�Ƃ��ĕK�v�Ȗʐς��傫���A���̍s�����͐l�Ԃ̐�����Əd�Ȃ荇�������������B���̂��߁A�l�Ԃ̊����������n�̔j��̌`�Ŕނ������������A�t�ɓ����̌̐��̑��������̊g�傪�_�ыƏ�̔�Q�������N�����肷��B���������āA�̐��̒��߂��܂ޓI�m�ȊǗ���@���m������Ȃ���A���̋�襂ȍ��y�ɂ����Đl�ԂƂ����̖쐶�������������邱�Ƃ͍���ł��낤�B
�{�����́A�쐶�����̕ی�Ǘ���@�m���̑����Ƃ��āA��L�W��̒��E��^�M���ނ̏�c�����A���͂����݂����̂ł���B
�䂪���ɐ������邨�悻120��̚M���ނ̂����A�W��݂̂�ΏۂƂ����̂́A��L�̗��R�̂ق��A���ɏq�ׂ钲�����@���K�p�\�Ȃ��̂Ɍ��������߂ł���B�Ȃ����R�����ΏۂƂ��Ă�����ׂ��J���V�J�iCapricornis�@crispus�j�́A���ɗގ��̒����i����1977�j���s���Ă��邽�ߏ��O�����B
�@
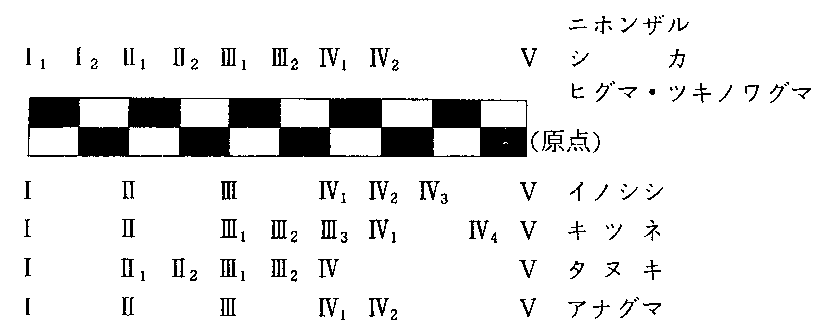
���i�P�j�@�{��͎��̂Q����ɋ�ʂ���Ă��邪�A�����ł͈���̋�ʂ͍s��Ȃ������B
�@�z���h�U���@�@M.fuscata�@fuscata
�@���N�V�}�U���@M.f.yakui
���i�Q�j�@�䂪���ɐ�������V�J�͊���̕ʈ��킠�邢�͕ʎ�ɋ敪����Ă��邪�A�����ł͐��ԓI�ɍ��̂���G�]�V�J�ƃz���V���E�W�J�݂̂���ʂ����B
���i�R�j�@�{��͎��̂Q����ɋ�ʂ���Ă��邪�A�����ł͈���̋�ʂ͍s��Ȃ������i�ʎ�Ƃ����������j�B
�@�j�z���C�m�V�V�@Sus�@scrofa�@leucomytax
�@�����E�L���E�C�m�V�V�@S.s.riukiuamus
���i�S�j�@��R���n����ɂ���ē����郁�b�V���i����b�V���A�Pkm���b�V���j�i�Q�|�P�Q�Ɓj���c�����ꂼ��T�{���ē����郁�b�V���ŁA����������Α���i�P/2.5���n�`�}�ɑ����j���c���Q�����������̂ł���B
�@
�����́A���a53�N�x�ɑS��47�s���{���S��ɂ����邻�ꂼ��̚M���ނ̐����n��A�����A���z�̕ϓ��i��Œn��A�o���N��A��ŔN��j�ɂ��āA�e�s���{���̒��b�ی���A�ޖ��W�E���Ȃǂ��A��҂Ȃǂ�ΏۂɁA�T�����̂P�n�`�}��16�����̂P��������ɂQ���������̂��ꂼ��Q�����ŁA�A���P�[�g�����[�ɂ�蕷���Ƃ���s�����i�����Ғ����v�ԎQ�Ɓj�B
�����l����2,235���A�����Ƃ�ΏێҐ��́A44,853���ł���B�����Ƃ���s�������͂T�����̂P�n�`�}��16�����̂P��摍����95.9%�ł���A��20���̏����ꂽ�B
�����Ώێ�̕��z�}�́A�e�s���{�����ƂɂT�����̂P�n�`�}��16�����������i�T�{�n�惁�b�V���A��Tkm�~�Tkm���b�V���F�ȉ��Tkm���b�V���Ƃ������i�S�j�ɂ���Ď�����Ă��邪�A���z���}�Ƃ��ẮA�T�����̂P�n�`�}��ɒ��ڐ����n�_���L�����ꂽ���̂��쐻���ꂽ�B
�@
���i�S�j�@�u�Tkm���b�V���v�Ɓu�Pkm���b�V���v�ɂ�镪�z�\����̊W
�{�����̕����Ƃ�n�_�́u�Tkm���b�V���v�łS�_�������ł���A������u�Pkm���b�V���v������Ɍ��Z�����0.16�_�ɂ����Ȃ��B����ɁA�R�x�n�тȂǂ̐l�Ƃ̂Ȃ��n����l����ƁA���̊����͂���ɉ����邱�Ƃ͓��R�ł���B���������āA�u�Pkm���b�V���v�\���ɂ�鐶����旦���́A�K�R�I�ɏ����Ȓl�ɂȂ炴��Ȃ��B
�܂��{�����̑ΏۂƂ����j�z���U���Ȃǂ̒��E��^�M���ނ̍s�����́A��������P�����L�����[�g�������傫���A�����̎�̐������n�_���Pkm�̏����ŕ\�������ꍇ�ɂ́A���̕��z��}�́A���ۂ����ߏ��ȕ\���ƂȂ�B���������āA�u�Pkm���b�V���v��P�ʂƂ��镪�z���ɂ�鐶����旦�������A�u�Tkm���b�V���v�ŕ\������������旦���̕����A���ꂼ��̎�̕��z���\������ꍇ�ɂ́A��莖���ɋ߂��Ƃ����悤�B
�u�Pkm���b�V���v�ɂ�镪�z���n�_�̏����́A��L�̂悤�ȕ��z��\����̌��ׂ͂�����̂́A��萳�m�ȕ��z�n�_��\�����Ă���̂ŁA��������傫�ȕ��z���̕\���ɕϊ����邱�Ƃ��ł���A���̍��y���l���̑����́A�u�Pkm���b�V���v�œ��͂���Ă���̂ŁA�����ƕ��z���Ƃ�ΏƂ����邱�Ƃ��ł���A�Ƃ������_�������Ă���B
�����ŁA���y���l���ƕ��z����ΏƂ�����ꍇ�ɂ́A�u�Pkm���b�V���v�\����p���邱�ƂƂ��A���z��̊g������d������K�v������ꍇ�ɂ́u�Tkm���b�V���v��p���邱�Ƃ������Ƃ��āA���z�v���̉�͂��s�����B
�@
�������ʂ����ƂɁA���a54�N�x�ɂ͊e�킲�Ƃ̓��{�S��ɂ킽��M���ނ̕��z�}���쐻���ꂽ���A���̍ی���������ł������`�F�b�N���邽�ߊe�������o���ꂽ���ʚM���ޕ��z���b�V���}�Ɗ��������Ɠ����̎�@�ŃA���P�[�g�������s�����M���ޕ��z�����Ȍ��O���[�v�ɂ�镪�z�������ʁi1977�N���{�j���Ƃ��r�������A�������ʂ̃Y���̂�����𒊏o�����̋��ɂ��čăA���P�[�g���s�����B
�Ȃ��A���z�}�̍쐬�Ɠ����ɂTkm���b�V���̋���P�ʂƂ���W�v�����Ƃōs���A���z�̑�܂��ȌX���̔c�������݂��B
����̒����͏�������܂����̓��e�������Ƃ蒲���[�ƕ��z���}�̂Q�ɂ킩��Ă��邽�߁A�W�v�E��͂ɂ������Ă͗��҂̏ƍ����K�v�ł����v�Z�ł͂قڕs�\�ł��邱�ƁA�܂����̕ۑ��������[���̂��̂̒����ۑ��������I�ɍ���ł��邱�Ɠ��̗��R�ɂ��A�����e�̎��C�e�[�v���͉����s�����B
���C�e�[�v�ɓ��͂��ꂽ�����e������̒����̌��f�[�^�|�̖������ʂ����ƂɂȂ邽�߁A���͍�Ƃɂ������Ă͒������e�A���z���}�L�ڂ̋L���n�_���\�Ȍ��蒉���ɓ��͂��邱�Ƃ�ڎw�����B���ɕ��z���}�ɋL�ڂ��ꂽ�n�_����͂���ɍۂ��Ă͒n�_���o���邾���������P�ʂœ��͂��邱�ƁA��̏W�v�E��͂ɂ������đ��̊֘A���Ƃ̂��荇�����\�ł��邱�Ɠ����l�����Ċ���b�V���i�O�����b�V���j��P�ʂƂ��ē��͂��邱�ƂƂ����B���z���}�ɋL�ڂ��ꂽ�n�_�͂��̊���b�V���ɂ��R�[�h���ɏ\���ς����鐸�x������Ɣ��f���ꂽ�B
���a55�N�x�ɂ́A���C�e�[�v�ɓ��͂������z���̏W�v�A�������s���Ƌ��ɁA���y���l���Ƃ̏d�˂��킹���s���A�܂��A�C�ے��̐ϐ��M���ނ̕��z�ɉe�����y�ڂ��ƍl��������v���̎��������W�����i�T�j�A���z�v���̎킲�Ƃ̕��͂��s���A���M���ނ̕��z�Ƃ��ꂩ��݂����R���ۑS�ɂ�������̑����l�@�����݂��B
�@
���i�T�j�@�ϐ�Ɋւ��鎑���̎��W�Ɛ���
�T���A�V�J�A�C�m�V�V�̕��z�Ɛϐ�Ƃ̊W���������邽�߂ɁA���̐ϐ�[���z�����~������̐ϐ�������z�}���쐻�����B��ƂȂ�ϐ�[�ɂ́A�ϐႪ�e��ɗ^����e�����l�����āA�T���ɂ��Ă�150cm�A�V�J�ɂ��ẮA�k�C���ł�60cm�A�{�B�ł�50cm�A�C�m�V�V�ɂ��Ă�30cm�����ꂼ��̗p�����B�ϐ�������z�}�̍쐻�͈ȉ��̂悤�ɍs�����B
���a43�N�i1968�N�j���珺�a53�N�i1978�N�j�܂ł̓~��10�V�[�Y���ɂ��āA��ƂȂ�ϐ�[���z����ϐ�������A�e�s���{���C�ی����ɂ���Ē��ׂ��B���̍ہA�k�C���ɂ�����60cm�ȏ�A�{�B�ȓ�ɂ�����30cm�ȏ�A50cm�ȏ�̐ϐ�����Ɋւ��ẮA10�V�[�Y���̂����ʎZ�T�V�[�Y���ȏ�ϑ����s���Ă���n�_����A�W���A�ʒu�����l�����đI�яo����1,120�n�_�ɂ��Ē��ׂ��B�{�B�ȓ�ɂ�����150cm�ȏ�̐ϐ�Ɋւ��ẮA���������Ȃ����߁A�P�V�[�Y���ȏ�ϑ����s��ꂽ���ׂĂ̒n�_�i282�n�_�j�ɂ��Ď������B�Ȃ��A������������ꍇ�ł��A�ϐ�[����L�̊�l�ɒB���Ă��邩�ۂ����s���ȓ������T���ȉ��ł���A���̃V�[�Y���ɂ��Ă̎����𗘗p�����B
���ɁA�ϑ��V�[�Y�����Ɗ�l���z����ϐ�����̍��v����A�e�n�_�ɂ������~������̕��ϐϐ�������Z�o�����B�����āA����ꂽ���l��50�����̂P�n���}��̊ϑ��n�_�̈ʒu�ɋL�����A�n�`�A�W�������l�����Ȃ���A�k�C���ɂ��Ă�80���܂ŁA�{�B�ɂ��Ă�70���܂ŁA10�����̓��ϐ���������������B����ɂ����̐}�Ɋ�Â��āA�e�Tkm���b�V���̕��ϐϐ�������e�ϐ�[�ɂ���10���P�ʂŋ��߂��B���̂Ƃ��A�ЂƂ̂Tkm���b�V���������̐ϐ�����敪�ɂ܂�����ꍇ�ɂ́A�ł��ʐς̑傫���敪���邢�̓��b�V���̂قڒ����Ɉʒu����敪���̗p�����B�܂��A�l���A��B�ɂ��ẮA��ƂȂ�ϐ�[�ɒB���������10���ȏ�ɂȂ�n�悪�������Ƃ���A�ϐ�������z�}�͍쐻���Ȃ������B���ӓ��ׂ����l�ɏ��O�����B
�@
�@���a53�N�Ɏ��{���ꂽ�����ɂ���ē���ꂽ20�����ɋy�ԕ��z���Ƃ��̌�Q���N�ɓn��W�v�╪�͂̌��ʂƂɊ�Â������Ώۂł���W��̒��E��^�M���ނ̕��z�ɂ��ďq�ׂ�B
�@�����Ɏ�グ���W�v�y�ѕ��͌��ʂ̑啔���́A�u��Q�R���ۑS��b�����������z�������i�M���ށj�S���Łi1979�j�v�y�сu�����̂Q�i1980�j�v�̓��e�����ɂP���z�̌���A�Q���z���K�肷��v���̕��͌��ʁA�R���z��̕ϓ��Ɋւ��鍀�ڂɂ��đI�����ĕ҂������̂ł��邪�A�ꕔ�ɂ͊����̔��f�������Ă���B
�@�ꕔ�̕\��p�ꓙ�ɂ��Ă͗l���ꂷ�邽�߉��ς����B���A�����̒̕��Җ�����̈��p�ɂ��ẮA�啔�����ȗ����Ă���̂Ō��T�ɑk�y����K�v������ꍇ�͌������Q�Ƃ��ꂽ���B
�}�Q�|�R�|�P�́A�j�z���U���̒n���I���z�����b�V���}�Ŏ��������̂ł���B
�{��́A�k�C���A��邨��щ��ꌧ�������P�s�Q�{41���ɕ��z���Ă���B���Ȃ킿���z��̖k���͐X�����k�����i�k��41�K31'�j�ł���A����͉��v���i�k��30�K20'�j�ł���B�{�B�A�l���A��B�̎��ӓ��ׂł́A���v���A�W�H�G�A�������ɂ������z����B�L�����̋{���ɂ��������Ă��邪�A���R���z�ł͂Ȃ��A���������ړ��������̂ł���B
������搔�i�Q�ꐶ����搔�{�Q��Ƃ͔���ł��Ȃ������̏o����搔�j�́A3,904���Ɩ{�����̑Ώێ�̒��ŁA�q�O�}�A�c�L�m���O�}�ɂ��ŏ��Ȃ��A������旦�i�{�킪���R���z���Ȃ��k�C���A����������S��搔�ɑ��鐶����搔�j�́A31.6%�ł���i�t�\�P�j�B
������旦��n���ʂɂ݂�ƁA�����A�ߋE�A�l���A�����A��B�����ꂼ��53.7%�A44.3%�A41.�V%�A34.4%�A32.9%�ƑS�����ς܂��͂���ȏ�ƂȂ��Ă���B
�֓��A���k�͂��ꂼ��21.6%�A11.9%�ƑS�����ς�肩�Ȃ�Ⴍ�A���Ƃɓ��k�͑S���ōł��Ⴂ�B�֓����瓌�k�n���ɂ����ĕ��z��ɋ��������Ƃ́A���̂��Ƃ���Ă���B
�S���I�ȕ��z�X��������ƁA�����n���Ȑ��̕��z�悪�L����������^�̐������z�������Ă���Ƃ����悤�B
�{��̕��z�悪���̂悤�ȕ��z�X���������̂́A���k�n���ł͂��̊���ȋC�A��B�n���ł͐A�ѓ��̎��R�n��̊J�����A�傫�ȉe����^���Ă���̂ł͂Ȃ����A�Ɛ��������B�����ŁA�C��̎w�W�Ƃ��Đϐ�[���A�l�דI�v���̎w�W�Ƃ��ĐX�ї����Ƃ肠���A�{��̕��z�Ƃ̊W�͂����B�܂��A�{��͈����їтɓK�����Ă��Ȃ��Ƃ̎w�E������A�{��̕��z��ƈ����їт̕��z��Ƃ̊W���A���킹�ĕ��͂����B
�P�@150cm�ȏ�ϐ�[�n��ƃj�z���U���̕��z
�j�z���U���́A�q�g�ȊO�̗쒷�ނł͐��E�I���z�̖k���Ɉʒu���A�[���ϐ�n�тɂ��������邽�߁A���̊���n�K���̐����A���ԓI�@�\�ɂ��Ă͏]�����璍�ڂ���Ă����B�������A����܂Ńj�z���U���̑S���I�ȕ��z�ƑΉ������āA�C��I�A���ԓI�ȕ��z�v�������͂��ꂽ���Ƃ͂Ȃ������B�����ł̓j�z���U���̕��z��̖k���A�k��35.3�x�Ȗk�A���o136�K�ȓ��̒n���ΏۂƂ��āA150cm�ȏ�̐ϐ�����̋��ƃj�z���U���̐������Ƃ̊W�ׂ��B�ϐ�[150cm�ȏ�̒n��́A�����闠���{�^�C��̒n��Ƃ悭��v���邪�A���̒n����̊e�Tkm���b�V����斈��150cm�ȏ�ϐ�[�̈�~���v�����ׁA�����10�����ɂW�敪�����̂W�敪�̐ϐ���ƃj�z���U���̐������Ƃ̊W�������i�\�Q�|�R�|�P�j�B
�Ώےn��ɂ�����150cm�ȏ�ϐ�n��̋�搔��4453���i�j�z���U���̕��z����n��̑S���n��搔��36.54%�j�A�Q�ꐶ����搔��553���i�S���̌Q�ꐶ����搔��25.96%�j�A�����̐�����搔��374���i�S���̏����̐�����搔��21.07%�j�A��ŋ�搔��151���i�S���̐�ŋ�搔��5.19%�j�ł������B���̂��Ƃ��A�j�z���U���̐������͐ϐ�n��ɂ��Ⴂ�䗦�ŕ��z���Ă��邱�ƁA�����ɐ�ŋ��̊������������ƁA�������Ă���B���Ȃ킿�A�j�z���U���̕��z��́A�����{�^�C��̐ϐ�n�����������Ƃ����X���������Ă���B���̂��Ƃ́A150cm�ȏ�ϐ�[�̒n��̋�搔�ɑ���Q�ꐶ���A��搔�̊����i�Q�ꐶ����旦�j������A����ɖ��炩�ł���B���Ȃ킿�A�ϐ�n��̌Q�ꐶ����旦��12.42%�ƑS���̗��n��搔�ɑ���Q�ꐶ����搔�̊����A17.48%�����A������Ă���B�������A150cm�ȏ�ϐ�[�̓������������Ă��A���̒n��ɑ���Q�ꐶ����搔�̊������������Ȃ����Ƃ͒��ڂ����B����́A�����̐�����旦�ɂ��݂邱�Ƃ��ł���B
��L�̃j�z���U���̕��z��Ɛϐ�[�Ƃ̊W��n���I�ɖ��炩�ɂ��邽�߁A�Tkm���b�V���Őϐ�[�ƃj�z���U���̕��z�}�Ƃ��d�˂��킹�A�}�Q�|�R�|�Q�Ɏ������B�j�z���U���̌Q��̐�����́A150cm�ȏ�ϐ�[���N��50���ȏ�ɒB����킪���ł��ł���̐[���n��A���_�R�n�A�����A�іL�R�n�A�z��R�n����є�ˎR���Ȃǂɂ����Ȃ�L�����z���Ă����B
�j�z���U���̕��z�悪����n��ɍL�������闝�R�Ƃ��āA�i�C�j�j�z���U���́A�����I�A���ԓI�ɐϐ�ɓK�����Ă��邱�ƁA�i���j����n��͑��тɓK���Ȃ��̂ŁA�����̒n��͎��R�тƂ��Ďc��\���������A���ʓI�Ƀj�z���U���̐����n��ۑS�������ƁA�i�n�j�j�z���U���̌Q��̐������̂��鑽��n��͂�������傫�ȎR��ł���A���сA����̐l�דI�e���������ɂ����n��ł��邱�ƁA�Ȃǂ��l������B
�Q�@�����їт̕��z�ƃj�z���U���̕��z
�j�z���U����v�ȐH���͖ؖ{�A���ł��邪�A�����їтɐ�������j�z���U���ł��A���їїv�f�̎����I�D����Ƃ��납��A�j�z���U���͈����їтɓK�����Ă��Ȃ��Ƃ̎w�E������B
�A���}�i�x��i1968�j�ɏ㌴�����M�j����A�{�B�̈����їђn��i�����n���Ȗk�ɂ�����{�B�̎�v�ȎR�x�̂قڂ��ׂĂ��Y������j���ʂ��o���A�j�z���U���̕��z�}��ɏd�˂��킹�Č��������Ƃ���i�}�Q�|�R�|�Q�j�A26�����̈����їђn��̂����A22�����ł̓j�z���U���̌Q��̐������͂Ȃ����A�����Ă����ӂ̂킸���ȕ������߂�ɂ����Ȃ��B�іL�R�n�Ȗk�̓��k�n���̈����їтł́A�����x���Ζʂ̈ꕔ���O�Ƃ��āA�܂������j�z���U���̌Q��͐������Ă��Ȃ��B
�����їт��L�����Ă���ɂ�������炸�A�j�z���U���̌Q�ꐶ����悪�݂���̂́A�����n���̂S�͏��ł���i�O���R���A����R�A�ԐΎR���A�ؑ]�R���j�B�����̒n��́A�O���R���̊␛�R�i2295m�j���ӂ������ƁA���ׂ�150cm�ϐ�[�̒n��O�ł���B
�����їт̑��݂́A�j�z���U���̕��z�𐧌�����d�v�ȗv���̈�ł���B
���ɐϐ�n��ɂ����ẮA�قڊ��S�Ƀj�z���U���̕��z�𐧌�����A�ƌ��_�ł��悤�B
�R�@�X�ї��ƃj�z���U���̕��z
�j�z���U���̕��z�ɉe����^����l�דI�v���ɂ��ẮA�X�є��̂ɂ�鐶����h���̖�肪1970�N���莩�R�ی�̊ϓ_����Ƃ肠�����Ă����B�{�ł́A�l�דI�e���𑪂�w���Ƃ��āA��R���n����i�Pkm���b�V���j�ɐ�߂�X�іʐς̊������i�U�j�i�ȉ��X�ї��Ɨ����j�ƁA�j�z���U���̕��z��Ƃ̊W�͂����B���͌��ʂ̈�Ƃ��ĐX�ї����S�敪���\�Q�|�R�|�Q�ɁA�e�X�ї��敪�ɂ����鐶����搔��Ivlev�̑I���W����n���ʂɎ����B���̏ꍇ�Pkm���b�V���\���̍��y���l���ł���X�ї��敪�̒n���I���z�ƃR���s���[�^���g�p���đΏƂ��邽�ߖ{��̐��������Pkm���b�V���ŏ����������̂�p�����B
�X�ї��敪�X�ɂ�����j�z���U���̐�����搔�͑S����9150���ł���A�X�ї�������Ƌ��ɐ�����搔�����Ȃ��Ȃ�X���́A�e�n���Ƃ����l�ł������B�܂��X�ы�搔�ɑ��鐶����搔�̊����i������旦�j���A��ʂɐX�ї�������Ə������Ȃ�X�����������B
���̒l�́A�e�X�ї��敪�̖ʐςɉe�������̂ŁAIvlev�̑I���W�������ȉ��̎��ɂ���������{��̐X�ї��I��x�Ƃ����B���̎���r�͂���X�ї��敪�ɏd�Ȃ�����̐�����搔�̂��̎�̑S������搔�ɑ��銄���i�\�Q�|�R�|�Q��B�j�Ƃ��AN�͊e�X�ы敪����������搔�ɑ��Đ�߂銄���i�\�Q�|�R�|�Q��A�j�Ƃ����
|
�X�ї��I��x�� |
r�|N r�{N |
�iIvlev�̑I���W���j
�Ŏ������B�����A����X�ї��敪��S�����p���Ȃ���i��������j���̒l�́|�P�ƂȂ�A���̐X�ї��敪���藘�p���Ă���P�ƂȂ�BO�̏ꍇ�ɂ́A�I�����Ă��Ȃ��i���W�ɐ�������j�Ƃ������ƂɂȂ�B
����ɂ��ƁA�X�ї��K��������ƑI���W��������Ƃ����͂����肵���X��������ꂽ�B
�ȏ�̂��Ƃ���A�j�z���U���̕��z�Ɛ�ρA�A���̊W�͎��̂悤�ɂ܂Ƃ߂邱�Ƃ��ł���B
���i�U�j�X�@�с@��
�M���ޕ��z���Əd�˂��킹��ׂ����y���l���Ƃ��āA�y�n���p�敪���̗p���ꂽ�B����́A�y�n�̗��p��Ԃ��A�X�сA�_�k�n�A�s�X�n�ɕ��ނ��A��O���n����i1km���b�V���j�̂��ꂼ��ɂ��āA�����̓y�n���p�����ꂼ��ǂ�ʂ̊����Ŋ܂܂�Ă��邩�����Ƃ��Ď�����Ă���B�X�т̎�ށi�l�H�т��V�R�т��j��X�т̑g���i�\����̓X�M���A�u�i���A�Ȃǁj�́A���̏��ł͔��ʂł��Ȃ��B�������A�Pkm���b�V���ɂ�����X�іʐς̊��������炩�ɂȂ�Ƃ������Ƃ́A�X�шȊO�̐l�ׂ̉e�����ɂ���y�n���p�����t�Ɏ������ƂɂȂ�B���������ĐX�ї����A���ΓI�Ȑl�דI�e���̓x���������w�W�Ƃ��Ďg�p�\�ł��낤�ƍl���A������S�i�K�ɂ킯�A���ꂼ��̋敪�ɁA�e��̐�����悪�ǂ�قNJ܂܂�邩�ׁA�W�v����Ƌ��ɁA�Qkm���b�V���̐}���쐬�����B
�{�ɂ�����e��̕��z�Ɗ��v���̉�͂Ɏg�p�����e�X�ї��敪�́A�ȉ��̂悤�Ɋe���ɑ���X�іʐς̊����������Ă���B
�X�ї��敪�X:�X�іʐ�70%�ȏ�
�X�ї��敪�U:�X�іʐ�40�`70%����
�X�ї��敪�R:�X�іʐ�10�`40%����
�X�ї��敪���̑�:�X�іʐ�10%����
�P�@����n��i150cm�ϐ�[����50���ȏ�̒n��j�̓j�z���U���̕��z�ɒ��ڂ̐�����^���Ȃ��B�������A����n��ň����їт�����ꍇ�́A�j�z���U���̕��z�͐��������i��O�A�����x���ΖʁA�␛�R���Ӂj�B
�Q�@�����їт͖��炩�Ƀj�z���U���̕��z�̐����v���̈�ł����āA�[���ϐႪ�݂��Ȃ��n��ł��j�z���U���̕��z�𐧌�����i��F�k��R�n�A�x�m�R�A���P�x�A��ԎR�j�B
�R�@�[���ϐ�̂Ȃ��n��ł́A�����їтł��j�z���U���̕��z��������i��F�ԐΎR���A�ؑ]�R���A����R�j�B����͐[���ϐႪ�j�z���U���̕��z��������x�������铭���������Ƃ������Ƃ������ƂƂ��ɁA�����̒n��̑��̊��v���i���Ƃ��A�����їтƂ����Ă��k���A���k�n���̂��̂ɔ�ׂ�ƁA���̎�g�����قȂ�Ƃ������A����́A���邢�͋C���̍����Ȃǂ̋C���̗v���j�̑���n��Ƃ̑�����������̂Ƃ��l������B
�S�@�X�ї��Ɏ������l�דI�e���́A�j�z���U���̕��z�𐧌�����ő�̗v���ł���B�j�z���U���͐X�ї��̒Ⴂ���i�X�тR�j�ł������邪�A�����̒n��ł̐������͂���߂ĒႭ�A���̂悤�Ȋ��́A�j�z���U���̕��z���m���ɐ������Ă���B
�j�z���U���̒n��̌Q�̑傫���������ڈ��ƂȂ�̂́A�Q�ꐶ����旦�i���n��搔�ɑ���Q�ꐶ����搔�̊����j�ł���A�ƍl������B�Q�ꐶ����旦�͓��k�n���ōł��Ⴍ�A��B�n��������ɂ��ŒႢ�B���n���ł́A�Q�ꐶ����搔�����Ȃ��̂ŁA�j�z���U���̕ی�㗯�ӂ��ׂ��n���Ƃ��čl������B���Ƃɓ��k�n���ł́A�l�ׂ̉e�����ɂ������R���A�j�z���U���̕��z�𐧌����鈟���їтƂȂ��Ă��邽�߁A�j�z���U���̑����͏�Ɋ댯�ɂ��炳��Ă���ƌ����悤�B���k�n���̃j�z���U���̕��z��́A�P�ɗ쒷�ނ̐��E�I���z�̖k���Ƃ�������łȂ��A���̐��ԓI�K���̋Ɍ����������̂Ƃ��āA���ɏd�v�Ȉʒu���߂���̂ƍl������B���k�n���ɂ�����j�z���U���̕��z��̕ۑS�́A�{��̕ی�ɂƂ��ē��ɏd�v�ł���B
�A�D���z�̌���
�}�Q�|�R�|�R�́A�V�J�̒n���I���z�����b�V���}�����������̂ł���B
�{��́A�k�C���A�{�B�A�l���A��B�A���˓������A�Γ��A�ܓ��A��������A�c�NJԏ����ɕ��z���邱�Ƃ��킩��B
�������Ȃ���A������搔�i���N������搔�{�ꎞ�I������搔�j��4,089���Ɩ{�����̑Ώێ�̒��ŁA�j�z���U���A�q�O�}�E�c�L�m���O�}�ɂ��ŏ��Ȃ��A������旦�i�S��搔�ɑ��鐶����搔�j�́A25.4%�ɂ����Ȃ��B�i�t�\�Q�j�B
������搔�͖k�C����l,684����M���ɁA�ߋE690.5���A��B690.5���A����511.0���̏��ł���B���k��65.0���͋ɂ߂ĒႢ�Ƃ�����B
������旦���݂�ƁA�k�C���A�l���A��B�����ꂼ��45.3%�A26.0%�A26.6%�ƑS�����ς܂��͂���ȏ�ƂȂ��Ă���B�{�B�ɂ��ẮA�ߋE��50.5%�ƍ������Ƃ������ẮA���k�A�֓��A�����A�����Ƃ��S�����ς��͂邩�ɉ�����Ă���B�Ƃ��ɓ��k��2.4%�Ƌɒ[�ɒႢ�B���z���b�V���}�ɋ��̑������Ƃ��A�����@���Ɏ����Ă���B�܂������ɂ��ẮA�k���n���ŋ��ڗ����Ă���B
�k�C���ƋߋE���A��搔�E��旦�Ƃ��ɍ����A�t�ɓ��k�E�֓��E�������Ⴂ�Ƃ�����B
�k�C���������ΐ�������^�̐������z�������Ă���B
�{��̐����͍���̒����ΏۂƂȂ����M���ނW��̒��ł̓j�z���U���ƂƂ��ɍň��̏�Ԃɂ���Ƃ����Ă悢�B���Ȃ킿�A������搔��4,089���i25.4%�j�ɂ������A���N������搔��1,963.5���i12.2%�j�Ƃ���ɂ킸���ł���B
�{��̌Q�̐��ޏ́A��搔�����łȂ��A���̕��z���n���I�ɐ��f����Ă���ɂ��݂邱�Ƃ��o����i�}�Q�|�R�|�R�y���\�Q�|�R�|�R�j�B���̂悤�Ȑ����搡�f�ɂ��n��̌Q�̎㏬���́A�����ȍ~�̒n��I��łɂ���Ă����i���ꂽ���Ƃ��l������B���̂悤�Ȑ�ŋ��W���n��͎���16�n��ł���B�P�j�n�����������@�Q�j�Ȗ،��ߐ{�E�����������́@�R�j�Q�n�������@�S�j�����s���k���@�T�j�É��������E���m�������@�U�j�������@�V�j�ΐ쌧�\�o�����@�W�j���䌧���E�k���@�X�j��㕽����ӕ��@10�j�L���E�����������@11�j�l�����암�@12�j�{�茧�암�@13�j�����������E���ꌧ�����@14�j���茧���˓��@15�j�ܓ��@16�j����������������B
���ʓI�ɂ́A�{��͓��k�n���ł͌ܗt�R�A���������E���؎R���������đS�����z�����A�����n��������n�т̖k���n���𒆐S�ɔ��n�悪�L�����Ă���B
�P�@�ϐ�ƃV�J�̕��z
�z���V���E�W�J
�z���V���E�W�J�iCervus�@nippon�@centralis�j�̊�������э̐H������ɂȂ�ϐ�[��45�p�Ȃ���50�p�ȏ�ł���Ƃ�����O�ώ@�̎�������{��̐����ɉe�����y�ڂ��ϐ�[��45cm�ȏ�Ȃ���50cm�ȏ�ϐ�������d�v�ł���ƍl������B����ɁA50cm�ȏ�ϐ������30�����z����Ɛϐ�ɂ�鎀�S�̂��o�͂��߁A50�����z����Ƒ�������Ƃ����������m�F����Ă���B�{�ł͂��̓_�ɂ��Ă���ɑS���I�K�͂Ō������邱�Ƃɂ����B
�\�Q�|�R�|�S�́A�{�B�E�l���E��B�ɂ�����50cm�ȏ�ϐ�����Ɩ{��̒ʔN������搔�E�G�ߓI�ꎞ�I�o����搔�E��ŋ�搔�Ƃ̑Ή����݂����̂ł���B
�ʔN������搔�́A50cm�ȏ�ϐ�[10�������ł͊��Ғl�ȏ�ƂȂ��Ă��邪�A�ϐ����10���ȏ�̋敪�ł͂���������Ғl�����������A���炩�ɒʔN������搔��50cm�ȏ�ϐ�[10�������敪�ɏW�����Ă���i92.2%�j10���ȏ�̋敪�ł͂���߂ď��Ȃ��A30���ȏ�̋敪�ł͂�������P%�����ƂȂ��Ă���B�G�ߓI�ꎞ�I�o�����́A��͂�ϐ����10�������ɏW�����Ă��邪�i86.6%�j�A10���ȏ�20�������敪�ł����Ғl���킸���ł͂��邪��܂���Ă���B�������A����ȏ�̓����敪�ɂȂ�Ɗ��Ғl�����܂��̂͒ʔN�������̏ꍇ�Ɠ��l�ł���B��ŋ���10���������ɏW������i85.5%�j�͓̂��l�ł��邪�A10���ȏ�20�������敪�Ȃ�т�20���ȏ�30�������敪�ł����Ғl���킸���ɏ�܂�鐔�l�������Ă���B30���ȏ�敪�ɂȂ��Ă͂��߂Ď��ےl�͊��Ғl�����������Ȃ��Ă���B
���̂悤�ɁA�z���V���E�W�J��50cm�ȏ�ϐ����10���ȏ�n��������X����ǂ݂Ƃ邱�Ƃ��o����B�G�ߓI�ꎞ�I�o����悪����n�тł��݂��邱�Ƃ́A�{���킪���U�E���Q�E�G�ߓI�ړ��Ƃ������ړ��l�����������Ƃ���l���Ă��蓾�邱�Ƃł���B
����50cm�ȏ�ϐ�[50���ȏ��{�B�ɂ�����z���V���E�W�J�̕��z�𐧌�����ϐ�ʂ̖ڈ��Ƃ��āA���̐ϐᕪ�z�Ɩ{��̒n���I���z�Ƃ̑Ή����݂��̂��}�Q�|�R�|�S�ł���B
�ʔN������悪���̊�ϐᕪ�z�ɐڂ��邠�邢�͂킸���ɏd�Ȃ�̂́A���Ɍ������A���k���A�ȖE�Q�n�������A���䌧�����A���쌧�k�������ł���B
�\�Q�|�R�|�T�́A50cm�ȏ�ϐ�[50���ȏ��搔�����ʂɂ݂����̂ł���B�H�c�E�R�`�E�V���̂R���́A50%�ȏ�̋�悪����Ɏ����ł��邪�A�����̌��ł͖{��͌������Ă��Ȃ��B
���������āA���̐ϐᕪ�z�͖{��̐���������ɂ��{�B�ɂ�����n���I���z�̍őO���ɔ�r�I�悭��v���Ă���Ƃ������Ƃ��o����B�{��̕��z�����̐ϐᕪ�z����u���Ă���ꍇ�́A�ϐ�ȊO�̕��z�����������Ă��ĕ��z�̋������Ă���Ƃ݂邱�Ƃ��o����B
�G�]�V�J
�Ƃ���Ŗk�C���ɐ�������G�]�V�J�iC.nippon�@yesoensis�j�̓z���V���E�W�J�iC.n.centralis�j�����ЂƂ܂��̂��傫���B���������āA�G�]�V�J�̏ꍇ�́A60cm�ȏ�ϐ�[�������ЂƂ̖ڈ��Ƃ��Đϐ�Ƃ̊W�ɂ��Č������邱�ƂƂ����i�\�Q�|�R�|�U�j�B
�G�]�V�J�̒ʔN�������́A30���ȏ�40�������ϐ������������80�������̂�����̐ϐ�����敪�ł����ےl�����Ғl����܂���Ă���B30���ȏ�40�������敪�ł����ےl�Ɗ��Ғl�̊u����͂킸���ł���B�ϐ�����敪80���ȏ�ɂȂ��Ă͂��߂Ċ��Ғl�������l���Q�{�ȏ����܂��A�G�]�V�J�����̐ϐ�敪������Ă��钥����݂邱�Ƃ��o����B�G�ߓI�ꎞ�I�o�����̏ꍇ�����l�ŁA80�������̂�����̐ϐ�����敪�ł����ےl�Ɗ��Ғl�̊Ԃɑ傫�Ȋu����͔F�߂��Ȃ��B�ϐ�����敪80���ȏ�ɂȂ��Ă͂��߂Ċ��Ғl�����ےl���1.5�{��܂��A�G�]�V�J�����̐ϐ�����敪������Ă��钥����݂邱�Ƃ��o����B
�}�Q�|�R�|�T�́A�G�]�V�J�̒n���I�敪�Ə�q��60cm�ȏ�80���ȏ�80���ȏ���̕��z�ƑΉ��W���݂����̂ł���B�ʔN������悪���̐ϐ�����敪�̕��z�Əd�Ȃ肠���Ă���̂́A���x�����E����E�x�ǖ�A�E�\���x�����E�͓��A�����R�����암�A�����x���W�ÂŔF�߂���B
�������A�����̏d���n��������A���҂͕��z��傫���قɂ��Ă���B���Ȃ킿�A60cm�ȏ�ϐ�[80���ȏ�̒n��̕��z�͖k�C���̐������ɕ��Ă���A����G�]�V�J�͂��̑���n��������悤�ɖk�C���̓������𒆐S�ɕ��z���Ă���B
�Q�@�X�ї��ƃV�J�̕��z
�\�Q�|�R�|�V�́A�n���ʂɂ݂��V�J�̐�����搔�ƐX�ї��敪�Ƃ̊W���݂����̂ł���B�Pkm���b�V���\���̍��y���l���ł���X�ї��敪�̒n���I���z�ƃR���s���[�^�|���g�p���đΏƂ��邽�߁A�{��̐��������Pkm���b�V���ŏ����������̂�p���A�e�X�ы敪�ɂ����鐶����搔��Ivlev�̑I���W�����������B
�X�ї��敪�X�ɂ�����V�J�̐������i�Pkm���b�V���j���́A�S����9,381���ł���X�ї�������Ƌ��ɐ�����搔���䂭�Ȃ�X���͊e�n���Ƃ����l�ł������B
�܂��X�ы�搔�ɑ��鐶����搔�̊������A�k�C�����������l�̌X�����������B����k�C���ł͐X�ї��敪�U�̂Ƃ���ɂ��̒l�̍ō����������B
�����Ivlev�̑I���W����p���ĕ\�킷�ƁA���̌X������薾�Ăɔc������A�z���V���E�W�J�́A�X�ї��敪�X��I��I�ɗ��p���A�G�]�V�J�͐X�ї��敪�U�����D�ށB
�ȏォ��A�V�J�͐X�ї��̍����n��ɏW�����Ă���Ƃ݂��邪�A�k�C���Ɠ��k�n���ȓ�ŌX�������قȂ�̂́A�y�n���p�̈Ⴂ�ɂ��̂ł͂Ȃ����ƍl������B���Ƃ��Ɩ{��͗щ����ł���̂ŁA�ނ���X�ї����ɒ[�ɍ����n������Ⴂ�n��̕����D�K�ł���̂�������Ȃ��B����ƁA�q��A����ȂǐX�шȊO�ł�������x�܂Ŗ{��̐��������e������y�n���p���s���Ă���k�C���ł́A����ڂ̐X�ї��敪��Ivlev�̑I���W���������Ȃ�͈̂ꉞ���Ȃ�����킯�ł���B����A�{�B�ȓ�ł́A�q��E���쓙�͏��Ȃ��A�_�k�n�E���Z�n�Ȃǖ{��̐��������e�����Ȃ��y�n���p���w��ǂł���̂ŁA���̂悤�Ȓn��ł̓V�J�̐����͐X�ї��̍����n��ɏW������̂ł͂Ȃ����ƍl������B
�R�@���
�}�Q�|�R�|�U�́A�u���b���v�v�ɂ��ƂÂ��吳12�N�i1922�N�j�ȗ��̃V�J�̕ߊl���̐��ڂ����������̂ł���B
�吳12�N�ȗ��A�ߊl���͑�Q�����E���O���������4,000���ȓ��őQ���X�����������A���ꂪ�}������̂͏��a25�N����37�N�܂ł̊Ԃł���B���a37�N�ɂ�13,000���ɂ܂ł͂˂������Ă���B�Ȍ�A���̑����͍Ăъɂ₩�ɂȂ邪�A���a50�N���ɂ�15,000����O�シ��܂łɎ���A�Ăы}���̒������݂��Ă���B���̔w�i�ɂ́A�l�������A�o�ϐ����ɂƂ��Ȃ��Ď�Ґ��̑����Ȃǂ��l�����邪�A������ɂ���{��ւ̎���͑����������ꌈ���Č������Ă��Ȃ����ƁA���Ȃ킿�{��̐��������͂��̓_�ł͂܂��܂������̈�r��H���Ă��邱�ƁA���̈����͋ߔN�����x�I�ɓx�����߂Ă��邱�Ƃ��l������B
���{�ɂ�����V�J�̏o���͔�r�I�V�����E�����X��������ɂȂ��Ă���ƍl������̂ŁA�{���̐�����́A�E�����X���Ȍ㕪�������k�C�����͂��߂Ƃ����{�y�Ǝ��ӓ��ׂƂ������ƂɂȂ�B�B�A���n���Ȃǂɖ{�킪�������Ȃ��̂́A���ݖ{�킪�������Ă��鉮�v���A��q���A�ܓ��A�Δn�A���˓��C�����ɔ�ׂē��̐����u�����͂邩�ɌÂ����߂ł���B�{��̒n���I���R�I���z�̕ϓ��́A�C��̊��g�ɑΉ���������n�т̊g��k���ɑΉ����A�{�B�ł͓�k�����A�k�C���ł͓��������ɕ��z��̕ϓ��������Ă����ƍl������B
�{��̐����͍���̒����ΏۂƂȂ����M���ނW��̒��ł̓j�z���U���ƂƂ��ɍň��̏�Ԃɂ���Ƃ����Ă悢�B���Ȃ킿�A������搔��4,089���A25.4%�ɂ������A���N������搔��1,963.5���i12.2%�j�Ƃ���ɂ킸���ł���B
�V�J�̌Q�̐��ޏ́A��搔�����łȂ��A���̕��z���n���I�ɐ��f����Ă���ɂ��݂邱�Ƃ��ł���B�����A�n���I���z���S�Ƃł������ׂ��̌Q�����z���Ă���n��́A�\�Q�|�R�|�R�Ŏ��������N������悪��r�I�L��ɂ킽���ĘA�����ďW�����Ă���I�̒n��Ƃ݂��A���ׂĉǐ�n�тɈʒu���Ă���B���̒n��̐X�ѐ�����旦�A���N������旦�͂�����������l�������Ă��邪�A���ɓޗnj��A�a�̎R���𒆐S�ɂ����I�ɔ����쓌���͂��̉\�����ł��傫���BI�������n��ł͂���ɕC�G����W�����͌����炸�A���z�悪�Ǘ������`�ɂȂ��Ă���B
���̂悤�Ȑ����搡�f�ɂ��n��̌Q�̎㏬���́A�����ȍ~�̒n��I��łɂ݂�ꂽ�悤�ɁA������V���Ȓn��I��ł݂������Ƃ��\�z�����B
���Ȃ킿�A�{��͓��k�n���ł͌ܗt�R�A���������E���؎R���������đS�����z�����A�����n��������̖k���n���𒆐S�ɔ��n�悪�L�����Ă���B�����E�l���E��B�n���ł̕��z������߂đa��Ŕ����Ȃ��Ă���A�{��̐����̕s����ȏ�\�킵�Ă���B
���̂悤�ȏɂ���Ȃ���A�V�J�̕ߊl�����傫���݂ď㏸��r�̌X���ɂ��邱�Ƃ́A�{��̓K�ȕی�Ǘ��̓_����A��l��v������ł��낤�B
�A�D�c�L�m���O�}�̕��z��́A�{�B�ł͒����ȓ��̎R�x�n�тɕ�A��������ѓ��k�n���ɍL�����z���Ă���i�}�Q�|�R�|�V�j�B�ߋE�n���암�ƒ����n�������ɂ́A���̕��z��Ɗ��S�Ɋu�����ꂽ���z�悪�݂���B�l���ł̕��z�͊ŏ��K�͂Ȃ��̂��_�݂��A�ɐB���̏��͓����Ȃ������B��B�ł͊m���Ȑ������͓����Ă��Ȃ��B
�c�L�m���O�}�̑�������搔��3,585���Ńq�O�}�Ɏ����ŏ��Ȃ��A������旦�i�{�킪���R���z���Ȃ��k�C���������S��搔�ɑ��鐶����搔�j��29.0%�ł���i�t�\�R�j�B
������搔�́A���k�n�����ł������A1,442���A���ɒ����n����1,323.5���ŁA���̂Q�̒n�悾���őS��������77.7%�Ɠ��{�S�̂̐������̑啔�����߂�B���̑��́A�֓��i297���j�A�ߋE�i270.5���j�A�����i26���j�̏��ł���B
������旦�ł��A���k�n�����ł������i52.5%�j�A�����i47.8%�j�A�֓��i23.8%�j�A�ߋE�i19.8%�j�A�����i16.0%�j�A�l���i3.9%�j�̏��ƂȂ�B
��B�ł͂��łɐ�ł������̂Ƃ݂Ȃ����B
�e�n���ʂ̕��z�����ڂ����q�ׂ�ƈȉ��̂Ƃ���ł���B
���k�n���F���̒n���́A�������̐җŎR�n�𒆐S�ɗאڂ����ȎR�n�̂قƂ�ǂ̒n��ɂ킽���ĘA�����ĕ��z�悪�L�����Ă���_�ŁA���̒n���ɂ݂͂��Ȃ�������L���Ă���B�����A���H�R�������ɔ��_�R�n�A�k��R�n�A���x�R�n�A�o�H�R�n�A�����E�іL�R�n����ђ�ߎR�n�ȂǂɘA�����z���݂���ق����k�����ɂ����z����B
�ȏ�̂悤�Ɉꕔ�ł��s�A���ȕ��z�悪���݂�����̂́A���k�n���ɂ�����c�L�m���O�}�̕��z��͂قژA�����Ă���Ƃ݂Ȃ��Ă悢���낤�B
�֓��n���F���k�n���ɑ��������җŎR�n����т��̗אڎR�n�ł���O���R�n�A�z��R�n�A�֓��R�n�ɘA�����z�悪�݂���B�u�����ꂽ���z��݂͂��Ȃ����̂́A���A��t�����̂��Ƃ��S��������悪�Ȃ����A������旦���Ⴂ�s�����܂ތ��ʁA�S�̂Ƃ��Ă̕��z��̍L����͂��قǑ傫���͂Ȃ��B
�����n���F���̒n���̕��z��͑O�̓��k����ъ֓��̂���ɗאڂ��A���A�����Ă���B��k���A���v�X�A�����A���v�X�͂��߁A��ˎR�n�A�����R�n�Ȃǂ���ȕ��z��ƂȂ��Ă���B
���̒n���ɂ͒���A�A����e���̂悤�ɁA���k�n���ɕC�G����L����������z�悪���݂���B
�ߋE�n���F�����R�n�ɗאڂ��Ĕ�ǎR�n��O�g�R�n�ȂǓ��{�C���ɖʂ����R�n�ɘA�����镪�z�悪���邪�A�����n���ȓ��Ɋr�ׂ��̍L����͒������������̂ƂȂ��Ă���B�Ȃ��A���̕��z��͓��k�n������̘A�����z�̐����ƂȂ��Ă���B�܂��A���̂ق��I�ɎR�n�ɁA���̂ǂ̕��z��Ƃ��q����Ȃ��u�����z�悪���݂���̂������n���ȓ��ł݂͂��Ȃ����ƂŁA���̒n���ł̃c�L�m���O�}�̕��z������Â��Ă���B
�����n���F�ߋE�n�������̕��z�悪�����R�n�����܂ŘA�����Ă��邪�A���R�A���您��ѕ��ɂ̌����t�߂܂łŁA�����萼�ɂ͍L�����Ă��Ȃ��B���������āA���̒n���̎����I�ȕ��z��́A�����R�n�����Ɉʒu���銥�R�R�n���ӂɂ݂��邾���ł���A�I�ɎR�n���l�A�u�����ꂽ�Ǘ����z��ƂȂ��Ă���B
�l���n���F��k�ɓ��ē����ɑ���l���R�n�̓����Ɛ����ɂ킸����������悪�_�݂���݂̂ŁA����܂ł݂Ă���������̒n���ɂ��Ȃ���������߂ĕn��ȕ��z��������Ă���B
��B�n���F���̒n���ł͕��z��͌��������A���łɐ�ł����Ƃ݂Ă悢�B�Pkm���b�V���ł͋{�茧���ɂS���݂̂��݂�ꂽ���A������������Ɋւ����N��Ȃǂ������Ă���A�����I�ɂ͐������Ƃ݂Ȃ����������̂ł���B
�C�D�c�L�m���O�}�̕��z�ƃu�i�ю��R��
����܂ł݂Ă����n���ʂ̃c�L�m���O�}�̕��z�́A�ꌩ���āA�{�B�A�l������ы�B�ɂ����闎�t�L�t���т̐����I���z�Ƃ悭��v�������̂ɂȂ��Ă���B�}�Q�|�R�|�W�̉䂪���̐����I�X�ёт̕��z�}�i���c1980�ɂ��j�ƃc�L�m���O�}�̕��z�i�}�Q�|�R�|�V�j���d�˂Ă݂���̑Ή��͖��炩�ł���B
�܂��A���t�L�t���тɐڂ����R�i���A�N���A�V�f�ށA���~�A�c�K�Ȃǂ���Ȃ�X�ёсi�g�ї��t���ёсj�������n�����瓌�k�n���̓������Ɍ������邱�Ƃ����킹��ƁA����痼�X�ётŃc�L�m���O�}�̕��z�͈�w�悭�J�o�[���邱�Ƃ��ł���B�Ȃ��}�Q�|�R�|�W�ɂ͈ܓx�ɂ����������I�X�ёт������Ɏ�����Ă��邪�A����́A��ɂ݂������{�Ńc�L�m���O�}�̕��z�悪�����ɂȂ邱�Ƃ́A�����ꏊ�Ƃ��Ă̗��t�L�t���ёт̋������ƌW���Ă��邱�Ƃ��悭�������̂ł���B
�Ƃ���ŁA�n���ʂ���ѓs���{���ʂɃu�i�A�~�Y�i���̐X�ёіʐςƃN�}�̕��z�ʐς̊W���݂��̂��\�Q�|�R�|�X���}�Q�|�R�|�X�ł���B�X�ёіʐς͑��R���ۑS�����A�������̐��ʁi�����A1976�j�ɂ��A�~�Y�i���|�u�i�N���X��̎��R����ё㏞�A���i�ȉ��ł̓u�i�тƗ��́j�ɋ敪���ꂽ�Pkm���b�V���̋�搔��S�Ă̌Q���ɂ��đ��v���邱�Ƃŋ��߂��B
�n���ʂɂ݂�ƂPkm���b�V������搔�ɐ�߂�u�i�ы�搔����уc�L�m���O�}������搔�̊����͓���X���������A�u�i�ы�旦�̏��Ɛ�����旦�̂���Ƃ͓���ƂȂ��Ă���B���̂��Ƃ͗��҂̊Ԃɐ��̑��֊W���������邱�Ƃ�����������̂ł���B�����ŁA�s�{�����ƂɃu�i�ы�搔�ɑ��鐶����搔���v���b�g���Ă݂�ƁA�����悻�̌X���Ƃ��ď�̊W�������ꂽ�B����ɂ��A�T���ē����{�ŔZ�������{�Ŕ����c�L�m���O�}�̕��z�̃p�^�[�����A������������邱�Ƃ��ł���
�������Ȃ���A���z��̍L����Ɛ����ꏊ�Ƃ��Ẵu�i�т̍L����̊Ԃɂ����܂��Ȑ��̑��֊W�������o�����Ƃ͂����A�s�{���ʂɂ͂������A�\�Q�|�R�|�X�ɂ��������Ƃ���u�����z������u�i�ш�̋����ߋE�⒆���n�����A�L���A�����z��������k�⒆���n�������t�ɍ������ŕ��z���Ă���A���҂̊W���P���ɐ������Ă�����̂ł͂Ȃ����Ƃ��Ɏ����Ă���B�{����܂ޑ�^�M���ނ̑S�Ă����Ă͎��A�����킸��Ώۂł���A�܂����R�̐l�ׂɂ����ς͏��ɂ���Ă��܂��܂ł��邩��A�n��ɂ���Č��݂̃c�L�m���O�}�̕��z����l�łȂ����Ƃ͓��R�̂��Ƃł���A���z�͂�����ė��j�̏��Y�ł���ƍl������B���̂��Ƃ͂������A��ł݂��c�L�m���O�}�̕��z��ƃu�i�т̍L����̊Ԃɂ́A�{�킪�����ꏊ���u�i�тɋ��߂�䂦�Ƀp�������ȌX�����������邱�Ƃ�ے肷����̂ł͂Ȃ��B
�Ȃ��A�k�C���̓암�A�n�������ɂ��u�i�т��������邪�A�c�L�m���O�}�̕��z���݂Ȃ��̂́A�n�j�I�Ȕw�i�������ƂȂ��Ă���ƍl�����Ă���B
�c�L�m���O�}�̕��z�����͏�ɂ݂��Ƃ���ł��邪�A���̂������z��̊g�傪�ߎ��ɂȂ��Ĉ����������ꂽ�͂�����Ƃ�����͂�����̒n��ɂ����Ă��݂��Ȃ��悤�ł���B�t�ɁA�n��I�Ȑ�łɂ�蕪�z�悪���ł�����A�n��̌Q�̐�ł̉\�����������z��̑��݂�A�s�A���Ȋu�����z�悪������Ƃ������ލs�I�ȕ��z��̕ϓ������炩�ɂȂ����B
���̂ق��A����̒����ł͐�łɊւ��������Ă��Ȃ����A���R�����炩�Ă͕��z��ł������ƍl������n�悪������݂��邱�Ƃ��������ꂽ�B
�����̕��z��ɂ��Ĉȉ��ɂ��ڂ����q�ׂ�B
��łɂ�镪�z��̏����F��B�n�����������邱�Ƃ͊��ɏq�ׂ��B����̒����ł́A�Tkm���b�V���ő啪���͂R���ɂW���A�{�茧��4.5����36�����ꂼ���ŏ����ꂽ�B�����͋�B�R�n�̑c��E�X�R�n�Ƌ{�茧�����̎R�n�ɔ�r�I���肳��Ă���B�܂��A��ł̔N��Ƃ��ẮA�����A�吳�Ƃ����{�茧��66%�A�啪����50%�������B
����v�ɂ��A���a26�N�ɑ啪���łR���̕ߊl���L�^����ĈȌ�ߊl�͓r�₦�Ă���B�܂��A�吳12�N�ȍ~�啪���ł͂킸���ɔN�P�`�S���̋L�^���O��V�N�ƂтƂтɂ݂��邪�A�{�茧���܂߂đ����ɂ݂͂�ׂ��L�^�͂Ȃ��B���̂悤�ɁA��B�n���ł̓c�L�m���O�}�̕ߊl���͂����͏��ł������B
��B�R�n�ł́A��q����悤�ɐ����ꏊ�̍L���肪���n��ɔ�r���Ē��������������Ƃ���A�n��̌Q�̋K�͂������Ȃ��̂ł���A����Ό����I�Ȑ��i�����������z��Ƃ��đ��݂��Ă����ƌ����悤�B�����āA�����I�ɒn��̌Q����ł��������͏��a�ȑO��z�肵�č����x���Ȃ����̂Ǝv����B
��łɕp���Ă��镪�z���F���̃P�[�X�Ƃ��Ă͎l���n������������B�l���R�n�ɂ͓����ɓ��Đ������݂�����̂́A�Ƃ��ɐ������͋͏��ŁA�����������ɐ�ŏ������܂�����悪�����A�ɐB�𗠕t�����F���ł��邱�ƂȂǂ���A�̌Q�Ƃ��Ă̑�������Ԃ܂��ɂ���B
�������̌��R�n�ƍ��m�A���Q�����̗���������r����Ɛ�ŏ����܂�����悪��҂ő����B�܂��A��҂ł͐�����悪���ɎU�݂��邱�Ƃ���A�n��̌Q�Ƃ��Ă͌�҂̕�����菬�������̂Ɛ��@�ł���B���̂��Ƃ͗��n����܂ޓ������ƍ��m���ɂ�����ߔN�̕ߊl���ɂ�������Ă���B����v�ɂ��A���a53�N�܂ł�10�N�Ԃ̕��ϕߊl���́A����4.8�A���m1.6�ł���B
��B�n���Ɠ������A�����ꏊ�̍L���肪�����A�����I�ȕ��z��̐��i�������Ƃ���A��łɌq����₷���ɂ���Ƃ�����B
�u�����z��̏o���F�I�ɎR�n�ƒ����R�n�����ɂقړ��K�͂̕��z�悪���݂���B
��������ȍ~�̐�ŏ��悵������̒����ŁA���R�A���挧������̂Ƃ��������R�n�̒����ƋI�ɎR�n�Ɍq����鎭�A�z������э����̊e�R�n�ɂ͐�ŏ�Ȃ��B���������āA���u�����z��������炵�������̒n��̕��z��̏��ŁA�����ꂽ���z��ƂȂ����͖̂����ȑO�̂��ƂƐ��@�����B
���u�����z�悪�o������Ɏ���ߒ��ɂ��ẮA�ꂽ���z��ɂ����鐶���ꏊ�Ƃ��Ă̐��ݐ��A���R���̐l���ρA����Ȃǂ��猟�����Ȃ���Ȃ�Ȃ����A�����������Ɏc�����ۑ�ł���B�����A�����R�n�ɂ��Ă͒n���w�̗��ꂩ��A�������ォ�狭���l�ׂ̂���ѕ��ɂ�莩�R���̉��ς������������Ƃ����A�c�L�m���O�}�̒n��I��łƂ̊֘A���ɂ����ċ�������w�E������B
�Ȃ��A���u�����z��ł́A��������ߔN�̉��n���R�т̐l�H���щ��i�g�呢�сj�ɂ�镪�z��̏k���ƕ��f�̐i�s����т���ɔ����n��̌Q�̎㏬�����F�߂��Ă���B
���Ȃ�Â�����̏��ŕ��z���F��B�R�n�̕��z�悪�����㏺�a�̏����ɏ��ł����̂ɑ��A���Ȃ��Ƃ������ȑO�ɒn��I�ɏ��ł����ƍl�����镪�z�悪����B
����̒����ł́A��ŏ�����Ȃ��������̂́A���k�n����ߋE����ђ����n���ŘA�����z��ɐڂ��A��Ƃ��ĐA������݂鎩�R�������ݓI�ɐ������\�ł���ƍl������n��ł́A���Ă̓c�L�m���O�}�����z���Ă����ɂ������Ȃ��B
�I�ɔ�������ђ����R�n�̐ꂽ���z��̑��݂ɂ��Ă͊��ɏq�ׂ����A���̂ق����k�n���ł͒Ìy�����ƈ������R�n�����̗�Ƃ��ċ�������B���������ł̌����Ǝ���͕s���ł��邪�A���̖����𖾂���ɂ́A�c�L�m���O�}�̐����j�͂������A�Ⴆ�Ήa�̗ʂ⎿�̔N�ϓ��Ɗ֘A�����̌Q���Ԃ����Ƃ̊W�Ȃǂ���������˂Ȃ�Ȃ��B
�ȏ�͔�r�I�L��ȕ��z��A�����n���I�ȕ��z�ɂ��Ă��̕ϓ����݂��̂ł��邪�A���z���n��̌Q��P�ʂɂ����ǒn�I�܂��͒n��I�ȕ��z���݂��ꍇ�A���z�̕ϓ��������Ă���P�[�X�������B
��Ƃ��Ď��R�т̔��̂Ƃ��̌�̐j�t���̐l�H���тɂ��c�L�m���O�}�̐����ꏊ�̊����ςړI�����Ƃ��āA���Ӓn��ւ̈ړ��A���U�������A�_�юY���ւ̉��Q�̑�����������ʁA�L�Q�b�Ƃ��ċ쏜�����Ƃ�����A�̉ߒ����A�n��̌Q�̕���Ɏ���p�^�[���Ƃ��Ēm���Ă���B
�A�D���z�̌���
�q�O�}�̕��z�͖k�C���{���Ɍ����A���ӂ̗����ɂ͐������Ă��Ȃ��i�}�Q�|�R�|�V�j�B
��Ȑ�����́A���A��ʁA�����A�_�U�Ȃǂ̎R�x�n�тɌ����A�s�s���A�_�n���̐i���암�ɂ݂͂��Ȃ��B
������搔�́A1,963.0���ƍ���̚M���ޕ��z�����Ώێ�̂Ȃ��ł��Ⴂ�l�������Ă���B�������A�{�킪���z����k�C���̑S��搔�ɑ��鐶����搔�A�ɐB��搔�̔䗦�A���Ȃ킿������旦�A�ɐB��旦�́A���ꂼ��52.8%�A20.5%�ƃc�L�m���O�}�̕��ϓI�Ȃ�����������A������旦�ł͓��k�n���̂��̂Ƌ߂��l�������Ă���B
��ȕ��z��͕X�I�Ɏ��̂U�n��ɕ����邱�Ƃ��ł���i�}�Q�|�R�|10�j�B�T.�������ȓ�̓n�������R�x��B�U.�ό���������x�܌��ւēo�ʎs�A�ɒB�s�Ɏ���n��̎R�x��B�V.���юR�n����V���R�n�ɂ����Ă̖k�����{�C���R�x��B�W.�@�J����I�z�[�c�N�C���̖k���R�n��ʂ���R���܂ł̒n��B�X.�[���R�n����ѓ����R����B�Y.�Y�y���爢���A�m���Ɏ���R�x�ȉ��ɂ��̂U�n��̕��z�ɂ��ďq�ׂ�B
�T��ł́A���ɓ��{�C���ɕ��G�Ȓn�`�����R�x�������A����炪�C�߂��܂ŕ��z���Ă��邽�߁A�N�}�̐�����������ɍL�����z���Ă���B���Y�p���͑�������J���Ă���A���ɒ������|���_�t�߂̊C�݈�ł͐�����قƂ�ǂȂ��B�T�c�������ɂ��܂��N�}�͐������Ă��邪�A���O�����ɔ�ׂ�Ə��ʂ͏��Ȃ��B�܂��A�������������̋�P�x���ӂł̓q�O�}�̐������͂Ȃ��B
�T��ƇU��̊ԁA���Ȃ킿�A��������n�тƃj�Z�R�A�R�A�r���R�A��������Ԑ��ɂ����܂ꂽ�y�ʊx�A�y���R�A���z�x���܂ށA��20�|50km�̒n��ł́A�q�O�}�̐�����قƂ�ǂȂ��A���݂ł͂��̒n��ɂ����Ď����㕪�z�����f����Ă���Ƃ݂邱�Ƃ��ł���B�܂��A�j�Z�R�A�R�A�r���R�ɐ�������Ƃ���鏬�̌Q���A���ӈ�̊J���ɂ��قڌǗ������Ă�����̂Ǝv����B
�U��̐�����ł͎x���Ύ��ӂɕ��z�̒��S������B���̐�����ł͐ϒO������œS���ƍ����T�������A�܂��A�������ł͎D�y�|��������ԍ���230���������f���Ă���A���Â����ʗʂ��������߁A�������̒n��ł̕��z�̕��f�ɂȂ���\��������B�U��̓����͐Ύ��n�тŁA�����͐l�ɂ��y�n���p���x���������ߌ��݂ł͂قڊ��S�ɕ��z�f���Ă���A15�|20km�ȏ�̔��тɂȂ��Ă���B
�V��͔�r�I�܂Ƃ܂����R�x��ŁA��ʗʂ̑������f���H�����Ȃ����߁A�N�}�̕��z�����قڈ�l�Ɋg�����Ă���B���̒n��͓�͐Ύ땽��A���͋�m����A���~�n�A����~�n�Ȃǂɂ���ĕ��z����Ă���A�k���͓V���쉈���ɑ��鍑��40������S���������ꉞ�̋��E���Ȃ��Ă���B�������A�k���ɂ����Ă͒��쒬��y�����̖k�C����w���K�ѕt�߂ɂ����āA�W��Ƃ̌𗬂��m���Ă���A���̕t�߂ł͍��Ȃ�������암�قǖ��Ăȕ��z�̒f��͂Ȃ��B
�W�͔��ɍL��ȕ��z��ŁA�암�̑��R��͓����R���ɘA�����Ă���A�����ł͌�ʗʂ̑�������38�������ꉞ�̋��E�Ƃ������A�q�O�}�̕��z�͂܂����̕����̈ꕔ�ɂ����ĘA�����Ă�����̂Ǝv����B�܂��A�����̇X��Ƃ��A�������R�x��������Ă���̂ŁA�q�O�}�̕��z�͈ꕔ�ŘA�����Ă�����̂Ǝv����B���̋��ł̕��z���͂قڑS��Ɋg�����Ă��邪�A���R���������k���₻�̎��ӂɂ����đ傫�ȋ悪����B�������A����͐������Ȃ����Ƃ��Ӗ�����̂ł͂Ȃ��A���̌��@�ɂ����̂ł��낤�B
�X��͗[���R�n�Ɠ����R�n�𒆐S�Ƃ����r�I�܂Ƃ܂������z��ł���B�����R���җ����ɉ����ĕ��z�̋������݂��邪�A�������@�ɂ����̂ł���B���̋�̍Ŗk���Ɉʒu���鍑��12���A38���A237�����ɂ����܂ꂽ�O�p��ł́A���݂̂Ƃ���܂��������F�߂��Ă��邪�A���̌̌Q�͂��łɂقڕ��z�����Ă���A�𗬂�����Ƃ���Γ암�̂����A�x�ǖ�s�|��ԓ쒬�Ԃ�12�|13km�̋�Ԃɂ����Ȃ��Ǝv����B
�W��ł͔��f�u�ˁA�����A�Õʒ��̎R�x�n�𒆐S�Ƃ���n��ƒm�������ɍL�����z��������Ă���B�A���A�Η��x�A�C�ʕi���܂m�������̐җ��R�����ł͐������Ă���Ǝv����ɂ�������炸�A��@�̂��ߋn��ƂȂ��Ă���B���̋�ƇW��Ƃ̋��E�͕s���Ăł��邪�A����A���ʁA�u�ˁA�k���ɂ����đ���S������э���242�����ɉ��������܂��n��ɐ������̏��Ȃ�����������B���̋�̒��ɂ͌��ݖk���̃p�C���b�g�t�H���X�g�𒆐S�Ƃ���n��ɌǗ����̌Q�Ǝv������̂�����A����͐��k�����������n��̌Q�̍Ō�̐����c��ł��낤�B
�ȏ�̂Ƃ���A�k�C���̃q�O�}��v���z���U��ɕ����Ă݂�ƁA�T�A�U��͂��łɌǗ����Ă��邪�A���̂S��͂܂��ꕔ�Ō݂��Ɍ𗬈�������Ă���B��҂ɂ����Ă��A���ꂼ�ꋫ�E��ɂ�����y�n���p���i�s���Ă���A����͂����̒n��ɂ����镪�z�̕��f�͂�薾�ĂɂȂ��čs�����̂Ɨ\�z�����B�܂��A���̂悤�ȕ��z�̕��f�͂����Ɏ��������̂���łȂ��A���ꂼ��̋���ł��e�n�ŋN��ł��낤����A�����I�ɂ݂�A�����͂���ɕ��z�̍ו������N��\��������B
���ɐX�тƂ̊W�Ńq�O�}���݂邱�ƂƂ���B�܂��A�q�O�}�̎�v���z��Ƃ��ĕ������U�n��̐X�т̏��L�`�ԂƂ̊֘A�ɂ��Ă݂�ƁA���̂悤�ȓ������F�߂���B�k�C�����̑S�X�іʐ�5,629,804�w�N�^�[���i���a54�N���݁j�̂����A���L�сi��w���K�т��܂ށj��3,219,674�w�N�^�[���i57.2%�j���߂Ă���A�V���W��ł̓q�O�}��v���z��̈ꕔ�ɁA���L�т̊ԂɊ��荞�ތ`�œ��L�т���r�I�L���ʐς��߂Ă��邯��ǂ��A��ʂɊe��̎R�x�n�̒��S���ɍ��L�сA���̎��ӂ̒�R�тɓ��L�сi11.0%�j���܂ޖ��L�сi42.8%�j�����z���Ă���i�}�Q�|�R�|10�j�B�����Ċe����ɂ�����q�O�}�̎�v���z�悪�قڊ��S�ɍ��L�тƏd�Ȃ�Ƃ���ɑ傫�ȓ���������B���̈Ӗ�����A�k�C���ɂ�����q�O�}�̌Q�̎�v���́A�R�x�n�ɑ傫�Ȗʐς��߂鍑�L�тɂ���Ĉێ�����Ă���ƌ����Ă��ߌ��ł͂Ȃ��B
��������̌����I�̌Q�̕��z�n�ł���p�C���b�g�t�I���X�g���r�I�o���L�^�̑����k���s�k���̒�R�ш�͂���������L�тŐ�߂��Ă���n��ł���B����A���L�ю��ӕ��Ɉʒu���铹�L�т₻�̑��̖��L�ђn��ł͑S��������Ȃ����A�܂��͂���̏��Ȃ��n�悪�����B���̎�v�Ȃ��̂͋T�c�����AI��ƇU��̒��Ԓn�сA�\�����쓌���u�˒n�Ȃǂł���B
���ɁA�X�ї��ƃq�O�}�̕��z�ɂ��Č����̂��\�Q�|�R�|10�ł���B�������A�O�q�̂Ƃ���A�����Ɏ������Pkm���b�V���\���ł̓q�O�}�̕��z�ُ͈�ɏ��Ȃ��L�^����Ă���A���̂��߂̐X�іʐρi���b�V���j����̕��z�䗦���ُ�ɒႭ�Ȃ��Ă�����̂Ƃ݂Ȃ���Ȃ�Ȃ��B���������āA������旦�̏ꍇ�Ɠ��l�A���̃f�[�^�͂قƂ�ǖ��Ӗ��ɋ߂����̂Ǝv����B����ɑ��A�q�O�}�̕��z����ɂ�����X�ї��ƑΉ��������z�l���i�䗦�j�̕��́A�X�ї��̈Ⴂ�ɂ���ċL�^���ɑ傫�ȍ����Ȃ����̂Ɖ��肷��A�O�҂��͈�w�L�ӂȂ��̂Ƃ����悤�B����ɂ��ƁA�\�z�����Ƃ���A�X�ї��̍���70�|100%�̒��Ƀq�O�}�̕��z���80%�ȏオ�܂܂�A�X�ї�40%�ȏ�̒n��ɂȂ��93%�ȏ�̕��z�悪�܂܂��B�܂��AIvlev�̑I���W�������߂�ƁA�X�ї��X�ł̂ݐ��̒l�������A���͕��A���Ȃ킿��������X�����������B
���݂̃q�O�}�̐�����́A�X�ї�40%�ȏ�̐X�ш�ɂقڌ��肳��A���̎�v�����́A�S�X�іʐς�57%���߂鍑�L�тɂ���Ĉێ�����Ă��邱�Ƃ����炩�ƂȂ������A�k�C���̊J���́A�e�n�̕��암���܂߂đS�悪�q�O�}�̐�����ł��������Ƃ͋^���Ȃ��B���Ƃ��A���݂͎D�y�s�Ɋ܂܂�A���̖k�����s�X�n�ɗאڂ��Ă���u��n��ɂ����āA����11�N12��25���ɋN�����q�O�}�ɂ��Y�ď����P�������▾��18�N�ɎD�y�̓����ɏo�������N�}�����Ȃǂ����̂��Ƃ��悭������Ă���B���̌�A�Ύ땽��A��m����A���~�n�A�\������ȂǁA��v�ȕ���̔_�ƊJ�����s����Ƌ��ɁA�����̒n�悩��q�O�}�̐�����͎R�x���Ɍ�ނ����B
���̂悤�ɁA���암���R�т̐X�ъJ���̐i�s�ƂƂ��ɃN�}�̐�����͌�ނ������A����ł����a30�N��܂ł͑啔���̒�R�т�R�[�悪�A�s�K���I���p�ł͂��邪���Ȃ��Ƃ��q�O�}�̋G�ߓI������ł��������Ƃ͑����̎�����������Ă���B����͐Ύ땽����ӎR�[��Ɋւ��Ă���O�ł͂Ȃ��B
���a30�N��ɓ����Ă��瓹���̊J���͋}���ɐi�݁A33�N����̊g�呢�ьv��ɂ�鍑�L�т̑�ʐϔ��̂Ƒ��сA35�N����̌o�ύ��x�����������ɂ��A�����n���@�J�V������ɂ�����X�є��̂Ɨ��_�p��K�͑��n�����q�O�}�̐�������k���E��ނ����������Ƃ��Ă����邱�Ƃ��ł���B�����̒n��ł͂��̎����Ȍ�q�O�}�̐�ň悪�g����A���݂ł͂قƂ�ǐ������͂Ȃ��Ȃ��Ă���B����A���K�͂ł͂��邪�A�������������Ǝv����������B���̈�͍�������̖�ꖜ�w�N�^�[���ɂ����Ď��{���ꂽ�p�C���b�g�t�H���X�g�v��ł���B����͏��a32�N����10�N�Ԃɂ킽���āA���̒n��̒Ⴂ�u�˒n�ɃJ���}�c�̑呢�ђn����������̂ł���B���ьシ�ł�20�]�N�������A�J���}�c�т����т��đ�X�т��������ꂽ���ʁA���a30�N�㏉���ɂ͂قƂ�ǂ݂��Ȃ������q�O�}�̑��Ղ��A�ߔN�ł͕p�ɂɂ݂���悤�ɂȂ��Ă���B����Ɏ����Ⴊ�n���������ɂ����Ă��݂���Ƃ����B���̒n���ł́A��R�т̐��̑��ђn�����т������ƂƉ��n�̐X�є��̂Ȃǂɂ��A�ߔN�ł͒�R�тɂ�����q�O�}�̏o�����p�ɂɂȂ��Ă����Ƃ�����B
�ꕔ�Ɉȏ�̂悤�ȗ�͂�����̂́A30�N��ȍ~�̓��H�Ԃ̔��B�Ƃ��̎��ӈ�̊J���ɔ����A�q�O�}�̐����悪�Q�����A�Ǘ������i�s���Ă��邱�Ƃ͋^���Ȃ����ƂƎv����B�c�L�m���O�}�ɂ������邱�Ƃł��邪�q�O�}�́A�l�ԂƐڐG����Ί�Q��^����\���̍��������ł���A�l�Ԃ̊�����ɓ��荞�̂͂قڊm���ɗL�Q�b�Ƃ��āA�쏜�����B
������̌Ǘ����A���f���̐i�s�́A���̂悤�ȋ@��������A�n��I�Ȑ�ł��炩�ẴG�]�I�I�J�~�̂悤�Ɏ�̐�łɂ܂Ŏ���\�����܂�ł���B���̂悤�ȑ�^�����̕ی�Ǘ��ɂ́A�l�ԂƐڐG����@��̏��Ȃ��L��Ȓn��̑��݂��s���ł���B
�A�D���z�̌���
�}�Q�|�R�|11�́A�{�����ɂ��C�m�V�V�̒n���I���z�����b�V���}�Ŏ��������̂ł���B
�{��̕��z�͑傫��������{�ɕЊ���Ă���B�k�C���A���k�̑啔���A����A�ΐ�̗������������k���Ƃ��������k���{�ɂ͑S���������Ă��Ȃ��B�܂��A�֓��A���A���m�A�F�{���̕��암�ł͕��z���a�ł��邩�A���z���Ă��Ȃ����Ƃ��ڗ��B
���ӓ��ׂł̖{��̐�����͏��Ȃ��A�{�킪���z���铇�ׂƂ��Ă͕��Ɍ��W�H���A�쐼��������������B�쐼�����ɐ���������̂̓����E�L���E�C�m�V�V�ƌĂ�A���킠�邢�͓Ɨ���Ƃ��ċ�ʂ����������B���茧�ܓ��ł͑吳����ɐ�ł����Ƃ���������Ă���B
�C�m�V�V�̐�����旦�́A���k��7.5%�ɂ͂��܂�A�֓��A�����̏��ɍ����Ȃ�A�ߋE�Ɏ����čō��l75.1%�������B����A�ߋE�Ȑ��ł́A�����A�l���̏��ɏ��X�Ɍ������A��B�ł�47.1%�ɂ܂Œቺ����i�t�\�S�j�B
�P�@�C�m�V�V�̕��z�Ɛϐ�
��ʂɗL���ނ͌㎈���߂̍����ȏ�̐ϐ�ɂ���čs�������j�Q����A�C�m�V�V�ł͖�30cm�̐ϐ�[���s���j�Q�ɂȂ���ƍl�����Ă���̂ŁA�����ł͐ϐ�[30cm�ȏ�̓������Ƃ肠���邱�Ƃɂ����B
�\�Q�|�R�|11�ɂ��A�ϐ������10�������ł͑S����50%�ȏ�ɐ������F�߂��邪�A�ϐ�����������Ȃ�ɂ�Đ�����旦�͌������A�ϐ�����敪70���ȏ�ł͂킸����3.3%�i56��j�������ɂ����Ȃ��B����56���̈ʒu�Ɠ���́A���쌧�P���A�E�ΐ�E����̌����n��51���A���Ɍ��S��ƂȂ��Ă���B
�{��̐ϐ�ɑ��鎩�R���z�̌��E�́A�P�~����30cm�ȏ㕽�ϐϐ����70�����ꉞ�̖ڈ��Ƃ��邱�Ƃ��\�ƍl������B
�{��̐������A�X�E�H�c�E�R�`�E�V���E�x�R�̊e�����͂��߂Ƃ�������n�тŔF�߂��Ȃ��̂́A�܂��������R�I���z�����v���Ƃ��Ă̐ϐႪ�A��Ɍ������Ă��邽�߂ł��낤�B
�Q�@�C�m�V�V�̕��z�ƐX�ї�
�\�Q�|�R�|12�́A�X�ї��敪�ɑΉ�������摍���Ɛ�����搔�i�Pkm���b�V���j�ł���B
�S�����v���݂�ƁA�{��̑S�������̖�80%�i16,814���j���A�X�ї�70%�ȏ�́u�X�тX�v�Ɋ܂܂�A�X�ї����Ⴂ�قǐ�����搔�����Ȃ��Ȃ��Ă���B�X�ї�10�������̋敪�ł���u���̑��v�ł͂킸��0.6%�A124���ɐ������F�߂���ɉ߂��Ȃ��B
�n���ʂɂ݂Ă��A�u�X�тX�v�Ɋ܂܂�鐶�����̊����́A���k�n���Ƌ�B�n����60%�Ƃ��Ⴂ�ق��́A�������80%�ȏ�������A�X�ї����Ⴍ�Ȃ�قǐ�����搔�����Ȃ��Ȃ�X���͈�v���Ă���B
�����Ivlev�̑I���W���ɂ���Ă݂�ƁA���k�n���ł́u�X�тX�v�ɑ���I�D�x�͌����łȂ��A�u�X�тU�v�ɂ����ėǂ��I�D����Ă���B�֓�����l���܂ł̊e�n���ł́A�u�X�тX�v���I�D����A����ȉ��̋敪�ł͊��������X�����݂�ꂽ�B
����A��B�n���ł́A�u�X�тX�v�ɑ���I�D�x�͊֓��n���Ɏ����ō����A�u�X�тX�v�ɑ��Ă������X���͔F�߂��Ȃ������B
�ȏ�̌��ʂ���A�{��͖��炩�ɐX�ђn�т𒆐S�ɕ��z���Ă���Ƃ�����B
�Ƃ���ŏ]���A�{��̐������Ƃ��ẮA�X�сE�k��n�E�l�Ƃ����݂���ꏊ���������Ă��邪�A����͍���̕��͂Ɩ���������̂ł͂Ȃ��A����̒���������A�{��͊m���ɍk��n��s�X�n��������x���݂���n��ɂ��������邱�Ƃ��킩��B��������ł��̔䗦�������Ȃ�Ɩw��ǐ������Ȃ��Ȃ邱�Ƃ������ł���B
�\�Q�|�R�|13�́A�X�ы敪�u�X�v�u�U�v�Ɋ܂܂���搔�ɑ��鐶����旦���Ȃ킿�����O�a�����݂����̂ł���B
�����O�a���̕��ς͖k�C���������Čv�Z�����49.6%�ŁA�܂��܂����z����g�債����]�n�͂��肻�������A���łɂ݂��悤�ɁA�k���E���k�n���͐X�т̏�Ԃ��悭�Ă�����̂��߂ɐ������������A����n�т������ƖO�a���͂���ɍ����Ȃ�B�֓��n����42.5%�Ƃ��Ⴂ���A�V���E�x�R�E�ΐ�E����S���������������n����66.8%�A�l���n��58.8%�A��B�n��52.9%�A�����n��68.5%�ƑS�����ς��������A�ߋE�n����82.0%�Ƃ���߂č����l�������Ă���B
�����O�a��80%�ȏ�̖O�a���敪�T�̊Y���n��͂T�{���ŁA�ߋE�n�������E�O�d�E����̂R�{���A�l���n���ł͈��Q���A��B�n���ł͋{�茧�ƂȂ��Ă���B�����Ő����O�a����60%�ȏ�80%�����̋敪�U�������̂�21�{���ŁA�ߋE�E�����n���𒆐S�ɂ��ċ�B�E�l���E��������ъ֓��n���̈ꕔ�ɕ��z���Ă���B�O�a��40%�ȉ��̋敪�́A��q��������n�тł���k���E���k�n���ɕ��z����ƂƂ��ɁA�l���̍��쌧�A��B�̍���E����E����̊e���ƂȂ��Ă���i�}�Q�|�R�|12�j�B
����n�шȊO�̒n��Ő����O�a���������Ă��錴���Ƃ��Ă͌�q�����ɂ��ߊl�����ɍl������B
����n�т������A���̖O�a���̒Ⴂ�n��قǍ��コ��ɕ��z���g�債����킯�ł��邪�A�����ł����u�X�сv�ɂ͖{��ɂƂ��ĕK�炸�����D�K�łȂ����̂��܂܂�Ă��邩�璍�ӂ�v����B
�E�D�C�m�V�V�̌Q�̒n���I���z�\��
���R�I���z�����v���Ƃ��Ă̐ϐ�Ƃ̊W����݂�Ȃ�A�{��̕��z���S�͖��Ⴀ�邢�͉ǐ�n�тł���A����n�тւނ����ĕ��z���ӂ��g�����Ă���Ƃ݂邱�Ƃ��ł���B�������A�e��̊J�������j�I�ɓW�J���ꂻ���ďW�ς��Ă��鍡���A���R�I���z��������݂����z���S���A�l�דI���z�����v���������e�����Ȃ��n��ɐ��������Ƃ݂邱�Ƃ��ł���B���̂悤�Ȓn��́A���łɕ��͂����悤�ɂ܂������X�ы敪�u�X�v�Ȃ����u�U�v�̒n��ł���B�������A�����ł�������͂��߂Ƃ������z�����v���������Ă��邩��A���݂̕��z���S�͂����̕��z�����v���̉e���ɂ�������炸�������͂�L����̌Q���������Ă���n��Ƃ݂邱�Ƃ��o����B
��ɕ��͂����ΐX�ѐ����O�a���́A���̂悤�Ȍ̌Q�̐��͂𐄗ʂ���ЂƂ̖ڈ��ƍl������B���������āA���̐����O�a���������n�悱�����͂̋����̌Q�����z���Ă���\�����傫���A���Ȃ킿���z���S�ł���\�����傫���Ƃ݂邱�Ƃ��o����B
�}�Q�|�R�|12�Ŗ��炩�Ȃ悤�ɁA�{�B�ł͑ΐX�ѐ����O�a���̍����n��́A�ߋE�n���𒆐S�Ƃ��Ē����n���A�����n���ւƊg�����Ă��邩��A�����̒n��͕��z���S�Ƃ݂Ȃ�����B����A�k���A�֓��A���k�n���ł̖O�a���͂���߂ĒႭ�A�������Ȃ��n�悷��F�߂��邩��A�����̒n��͕��z���ӂƂ݂Ȃ��č��x���Ȃ��B����A�l���n���͈��Q���ł̖O�a��������߂č����A���m�E��������������Ɏ����A���쌧�ł�20%�ȉ��Ƃ���߂ĒႭ�Ȃ��Ă���B���������āA�l���ł́A���Q���𒆐S�Ƃ����n���I���z�\���������Ă���Ƃ݂���B��B�n���ł́A�{�茧�̖O�a��������߂č����A�אڂ���啪�A�������A�F�{���������ō����Ȃ��Ă���B�����ĕ����A����A���茧�̏��ɒႭ�Ȃ��Ă���B���R�̂��ƂȂ���A��B�n���ł͋{�茧�z���S�Ƃ��A�k�E����B�z���ӂƂ��镪�z�\���ƂȂ��Ă���B
�i�U�j�L�c�l�E�^�k�L�E�A�i�O�}�̕��z
�A�D���z�̌���
�P�@�L�c�l�̕��z
�}�Q�|�R�|13�́A�{�����ɂ��L�c�l�̒n���I���z�����b�V���}�Ŏ��������̂ł���B
�L�c�l�́A��B�A�l���A�{�B�A�k�C���ɂ���߂čL�����z���A������搔��9,781.5��悪��������B����̑S���ɑ��銄�����Ȃ킿������旦�͎���60.8%�ɂ��̂ڂ�A�^�k�L�ƂƂ��ɂ܂������S���I�ɕ��z���Ă��邱�Ƃ��킩��B�i�t�\�T�j�B�������A���ӓ��ׂł̐�����͏��Ȃ��A�킸���ɖk�C���̗��K���ƒ��茧�̌ܓ����m���Ă���ɂ����Ȃ��B���Ɍ��W�H���A�k�C���̗當���ł͂��ꂼ�ꏺ�a30�N��A40�N��ɐ�ł��Ă���B����ȊO�ɕ��z�̋��ڗ��̂́A�i�P�j�R�x�n�щ��n�i�Q�j���암�i�֓��A�z��A�������쓙�j�i�R�j�l���i�S�j���B�A�ł���B�R�x�n�щ��n�̏ꍇ�A���l�n�т������̂ŕ���������ꂸ�A�������Ă��Ă��m�߂��Ȃ��������Ƃ��l������B���암�ł͂��̂悤�Ȃ��Ƃ͖w��Ǎl����ꂸ�A���炩�̌����ɂ���Đ�ł��i�s�������ʂƂ݂���B�l���A���B�Ŗ{��̕��z������߂Ĕ����͓̂����I�ł���A���̌����Ƃ��ẮA��̂Q�_�ɗ��R�����邩������Ȃ��B
�Ƃ���Ŗ{��̐�ŋ��̑S�����v��762.0���ŁA4.7%�A�S�������ɑ��Ă�7.8%�ł������B
������旦�̍ō���78.9%�A�Œ�͎l����13.8%�ƂȂ��Ă���B��B��40.�V%�������A�l���Ɣ�ׂ�Ƃ͂邩�ɍ����l�ƂȂ��Ă��邪�A�����Ȗk�̒n����50%�O�゠�邢�͂���ȏ㎦���Ă���̂Ɣ�ׂ�Ƃނ���Ⴂ�l�ł��邱�Ƃ��킩��B
�Q�@�^�k�L�̕��z
�}�Q�|�R�|14�́A�^�k�L�̑S�����z�����b�V���}�Ŏ��������̂ł���B
�^�X�L�́A�k�C�������B�܂łقڑS���I�ɕ��z���A������搔��9,830.0���A������旦��61.1%�ƂȂ��Ă���i�t�\�U�j�B�������A���ӓ��ׂ̂����A���K���i�k�C���j�A���n�A�B��A���A���i���������j�A�V���㓇�����i�F�{���j�A�W�H���E�������Ȃǂ̏��Ȃ���ʐ��˓��C�̓��X�ɂ͕��z���邪�A�쐼�����A�ܓ��A�Δn�A�ɓ������A���K���A�當���Ȃǂɂ͕��z���Ă��Ȃ��B
�܂��A�k�C���ł́A���G�x���Ə@�J�x���̖k���ŕ��z����r�I�A�����Ă���̂������A�啔���̒n��͐�ꂠ�邢�͂��ȕ��z�ƂȂ��Ă���A�������̎R�x�n�тƖԑ��x���������獪���x���ɂ����ẮA�{��̕��z��H�ł���B
�{�B�Ő��������ꂸ�A���z�����邢�͂܂�Ȓn��́A���k�����A�Ìy����A�H�c���쓙�̓��k�n���k���A�z�㕽��A�������A�E��������сA�֓�����A�k�A���v�X�A��A���v�X�A�Z������A���Ð여�擙����������B�l���n���ł͐��˓��C���ݕ��̐V���l�A�]��A�����Ȃǂ̕��암�A�ΒȎR���̉��n�R�x���ŕ��z�����邢�͂܂�ɂȂ��Ă���B
�ȏ�̕��z���̂�����ŏ����ꂽ���͑S���ō��v378.0���A�S��搔�ɑ���2.4%�A�S������搔�ɑ��Ă�3.7%�Ɣ�r�I�����Ȓl�ƂȂ��Ă���B
�R�@�A�i�O�}�̕��z
�}�Q�|�R�|15�̓A�i�O�}�̒n���I���z�����b�V���}�Ŏ��������̂ł���B
�A�i�O�}�́A�{�B�A�l���A��B�قڑS��Ɛ��˓��C�����̈ꕔ�ɕ��z���A�k�C���A���n�A�ɓ������A�B��A���˓��C�����̑啔���A�듇�A�Γ��A�ܓ�����ё�������ȓ�̓쐼�����ɂ͕��z���Ă��Ȃ����Ƃ��킩��B�܂��A���������A�m�������A���������̔����ł̓A�i�O�}�͐������Ă��Ȃ����A���̂��Ƃ͗����ɃA�i�O�}�����Ȃ����ƂƂƂ��ɋ�����������ł���B����ɉ����ĉ��k�����A�Ìy�����A�\�o�����A�F�������A����������̔������ł��A�i�O�}�̕��z�͑e�ɂȂ��Ă���B����ɐ��A�V���A�����A���É��A���A�����A�F�{����s�s���ӂ̕��암�ŃA�i�O�}���݂�ꂸ�A��ň悪�������Ƃ������I�ł���B
�S���̐�����搔��5,876.5���ŁA�^�k�L�A�L�c�l�Ɏ����ő����A������旦�i���R���z���Ȃ��k�C�����������S��搔�ɑ��鐶����搔�j��47.5%�ł���i�t�\�V�j�B
�^�k�L�A�L�c�l�A�A�i�O�}�ɂ��Ė��炩�ƂȂ����_�����Ă݂�ƁA�P�j���암�ł͂R��Ƃ�������悪���Ȃ��A���R�n���������旦���Ⴉ�����i�R�x�n�т͐���������ɂ������Ƃɂ����j�B�Q�j�s�s���̐i�n���A���Ɋ֓��ɂ����ĂR��Ƃ���ŋ�旦�������Ȃ��Ă����B
����ɁA�L�c�l�͎l���A��B�ɂ����Đ�����旦���Ⴍ�A�^�k�L�͖k�C���ŒႩ�����B�A�i�O�}�͑��̂Q��ɔ�ׂ��Ⴂ������旦�������Ă���A�k�C���Ɏ��R���z���Ă��Ȃ��ق��́A�S�����l�Ȑ�����旦�ł������B�Ȃ��A����ɂ͂��̂R��Ƃ����R���z���Ă��Ȃ������B
�����R��̒��^�M���ނ́A�q�O�}�A�c�L�m���O�}�A�V�J�A�C�m�V�V���̑�^�b�̂悤�ɕ��z���݂��Ă��炸�i���Y��̕��z�}���Q�Ɓj�A�S���I�ɂقڈ�l�ȕ��z��Ԃ������Ă���B���̎��́A�ނ炪�G�H���ł��邱�ƂȂǂ���A�V�J�Ō�����ϐ�Ȃǂ̊��v���ɂ���Ă��قǂ��̕��z����������Ă��Ȃ����Ƃ��Ӗ�����ƍl������B����A���̐����悪�l���ɋ߂����Ƃ���A�l�Ԋ����̉e�����������ł���Ƃ�������ł��낤�B
�C�D�X�ї��ƃL�c�l�E�^�k�L�E�A�i�O�}�̕��z
���z�������C�e�[�v�w���[����ہA�R�����b�V���ilkm���b�V���j�ɂ�萶����搔�Ă��邪�A���̋�搔���T�~�Skm���b�V���œ���ꂽ������搔�Ŋ������l�́A�������̑e���������ƍl������B���Ȃ킿�A�T�~�Skm���b�V���Ő����Ƃ��ꂽ��悪�R�����b�V���ɕ������ꂽ���A�����̋�悪�������ł�����̒l�͍������̂ƂȂ�A����������Ȃ킿�������m���Ȃ��̂ƍl������B�܂��A���̒l�͐������x�����̂܂ܔ��f������̂ł͂Ȃ����A�������x��������Ώ�������Ȃ�ł��낤���番�z���a�ł���ΒႭ�A�W�����Ă�������X�������ł��낤�B
�L�c�l�A�^�k�L�A�A�i�O�}�ɂ��āA�S���̑�������搔�ŋ��߂��l�͂��ꂼ��A2.69�A3.20�A2.32�ł������B�}�Q�|�R�|16�ɂ͓s���{���ʂɋ��߂����̒l�̃q�X�g�O�������R�했�Ɏ����Ă���B�A�i�O�}�͂Q�����[�h�Ƃ��ĒႢ�l�Ɋe�����W�����Ă���A�S����l�ɑa�ȕ��z�����Ă���ƍl������B�L�c�l�A�^�k�L�͌����Ńo���c�L�������B
���ɁA�����R��̕��z�l�����������邽�߂ɁA�X�т����̔�x�ɂ��S�ɋ敪���A�������Ƃ̏d�ˍ��킹�������Ȃ����B�X�ї��̊e�敪���ɐ�����搔�����̎�̑S������搔�ɑ��Đ�߂�������\�Ɏ����Ă����B���̒l�́A�e�X�ї��敪�̖ʐςɉe�������̂ŁAIvlev�̑I���W�������߁A�X�ї��敪�ɑ���I�D�x���r�����B
�}�Q�|�R�|17�ɃL�c�l�A�^�k�L�A�A�i�O�}�ɂ��Ă��̒l���O���t�Ŏ����Ă���B�R��Ƃ����ɑI��I�ɗǂ����p����X�ы敪�݂͂��Ȃ����A�L�c�l�A�^�k�L�͐X�ѓx�U�i�X�ї���70�`40%�j�Ɏ�����l�������Ă���B�X�т��̑��i�X�ї�10%�ȉ��j�͂R��Ƃ��������Ă���A�i�O�}�œ��ɂ��̌X���������A�A�i�O�}�A�^�k�L�A�L�c�l�̏��ƂȂ��Ă���B���̌X������A�A�i�O�}�͐X�ї�40%�ȉ��̏��͐����K�n�ł͂Ȃ��ƍl�����A����A�L�c�l�͂��Ȃ�X�ї����Ⴍ�Ă���������ƌ����邾�낤�B�L�c�l�́A�k�n�A�q���n�Ȃǂł̊ώ@��������A�����œ���ꂽ���ʂ́A�ŋߎ���ɖ��炩�ɂ���Ă��邱���R��̐��Ԃ�����x�������B�����R��́A�T���Đl���ɋ߂��������铮���ł���Ƃ����悤�B
�L�c�l�A�^�k�L�̏ꍇ�A������旦�̍������͐�ŋ�旦���Ⴂ�X���ɂ������B�܂��A��ŋ�旦�̍������͊T���ēs�s���̐i��ł���n��ł������B���̂��Ƃ́A������旦�A��ŋ�旦�Ƃ��ɐX�ї��Ƃ��̖ʐςɖ��ڂɊ֘A���Ă��邱�Ƃ��������Ă���B
�\�Q�|�R�|14�ɂ͐�����旦�̍��������P�{�T���Ɛ�ŋ�旦�̍��������P�s�P�{�V���̂��ꂼ��̋�旦�ƐX�ї�10%�ȉ��̋�旦�������Ă���B
������旦�̍��������{���͐X�ї�10%�ȉ��̋�悪���Ȃ��B�܂��A�R�����b�V�����T�~�Skm���b�V���Ŋ������l�������A���Ȑ���������Ă���B�������ꂾ���͐�ŋ�旦�������B�X�ї�10%�ȉ��̋�旦�������A�Pkm���b�V���ƂTkm���b�V���̐�����搔���r�����l���������Ƃ���A���ꌧ�ł́A���암�ł͐�ŌX���ɂ�����̂̎R�n�ɂ����đ����̒n��Ő������F�߂��邱�Ƃ������Ă���̂ł��낤�B
��ŋ�旦�̍��������s�{���̐X�ї�10%�ȉ��̋�旦�͐�����̗ǂ����ɔ�����A��ʁA�����A���A��t�A�����̏��ƂȂ�B�����̒n��͕��암�̑������ƁA�����ēs�s�����i��ł��邽�߂ɐ������s�K�Ő�ŋ�旦�������Ȃ����ƍl������B��ʂ́A�R�����b�V���ƂT�~�Skm���b�V���Ő�����搔���r�����l�������Ȃ��Ă���B���̂��Ƃ́A�֓�����ł͐�ł̌X���������є\�|�{�������ԃ��C���Ȑ��ł͋t�ɐ������W�����āA������ɒn��I�ȕ݂����邱�Ƃ������Ă���B
�H�c�A�ΐ�A�a�̎R�A�{��̂S���͐X�т������ɂ�������炸��ŋ�旦�������B�ΐ�́A�L�c�l�̐�ŋ�旦�̒��ōł������l�������Ă���B
�L�c�l�Ɠ��l���\�Q�|�R�|15�Ƀ^�k�L�̐�����旦�ƐA���Ƃ̊֘A�������Ă���B
������旦�̍����e���́A�X�ы敪�g���̑��h�̐�߂銄�������Ȃ��A�R�����b�V���ƂT�~�Skm���b�V���̊������������̂ƂȂ��Ă���B���ꌧ�̓^�k�L�ɂ����Ă������Ԃ��ŋ�旦�������X���ɂ���A���암�őa�A�R�n�ŏW���������z�l�����Ƃ��Ă�����̂ƍl������B
��ŋ�旦�̍����s�{���͂����ł����암�������X���������Ă���A�^�k�L�̐�������܂��s�s���̐i�s�Ɩ��ڂȊ֘A������B�Ȃ��ł��A��ʁA�����A���̓L�c�l�Ɠ��l�ɂ��̌X���������ł���B
�A�i�O�}�͑O�q�����悤�ɃL�c�l�A�^�k�L�ɔ�א�����旦�A��ŋ�旦�Ƃ��ɒႢ�l�ɑ����̌����W�����Ă����B�\�Q�|�R�|16�ɂ͑O�Q��Ɠ��l�ɁA�X�т̋�旦�Ɛ����E��ŋ�旦�������Ă���B������旦�̍����n��͐X�ы敪�g���̑��h���Ⴂ�X���������Ă��邪�A���܂薾�m�ł͂Ȃ��B����A��ŋ�旦�̍����n��́A���암�̑������ł������B
�\�Q�|�R�|17�ɂ́A�L�c�l�A�^�k�L�A�A�i�O�}�ɂ��Đ����A��ŋ�旦�̒Ⴉ�����n��������Ă���B����̓L�c�l�A�^�k�L�A�A�i�O�}�Ƃ���Ȃ��O�q�����悤�Ɏ��R���z���Ă��Ȃ��B�k�C���̃A�i�O�}�ɂ��Ă����l�ł���B�k�C���̃^�k�L�������A�ǂ̌������̎�Ɋւ��Ă̏�����Ȃ��A���Ȃ�ȑO�����ŌX���ɂ����Č��݂Ɏ������̂��A���Ƃ��Ǝ��R���z�̓x�������Ȃ��̂����Âꂩ�ł��낤�B�l���̃L�c�l�̐������̏��Ȃ����]�ˎ���̂��̒n���̓s�s���ɂ����̂Ƃ����������B�܂��A���m�����̓L�c�l�����u�ɂ���ł̉\�������邱�Ƃ���������Ă��邪�A�L�c�l�Ɠ����C�k�Ȃł���^�k�L�̐�����旦�̒Ⴓ�����u�ƊW������ƍl�����A���̐����̕ω��ɒ��ӂ��K�v�ł��낤�B�k�C���̃^�k�L�̐������́A���G�A�@�J�x���ɏW�����Ă���A���R�A�����R���A�\������ŏ��Ȃ��B���̗��R�̈�ɂ͎R�x�n�ł̐������̕s�����l�����邪�A�����Ŏ�肠�����X�ї��ȊO�̗v���i�Ⴆ�A�n�`�A�n�j�A�ϐ�j�����z�l���ɑ傫���e�����y�ڂ��Ă��邱�Ƃ��l������B
�L�c�l�A�^�k�L�A�A�i�O�}�̐�ŋ�旦�̍��������n��ɂ��āA�N��ʂɓ���ꂽ��ŏ�炻�̉ߒ��̕��͂������Ȃ����B��ŏ��͖����A�吳�A��O�A��20�N�A��30�N�A��40�N�A��50�N�A�s���ɋ敪����Ă���i�t�\�W�`�X�j�B�N�㏇�ɏ��ݐς������ɑ���S���������߂Đ}�������B���̍ەs���̏�͑����̒��ɂ͊܂߂����ݐϐ��ɉ����Ă��Ȃ��B���������āA�O���t�̍Ō�̓_�i��50�N�j�́A100%�ɒB���Ă��Ȃ��B
�}�Q�|�R�|18�ɃL�c�l��ŋ�旦�̍��������n��ɂ��Ď����Ă���B�O���t�́A��ʁA��t�A�����A�{��̂悤�ɌX�������₩�Ŗ����̐�ŏ�������n��ƃO���t�̂���������̋}�Ȓn��ɕ�������B��ʁA��t�A�����Ȃǂ́A�s�s���ƂƂ��ɖ����ȑO����L�c�l�̐�ŌX�����������ƍl������B�}���̖��͐�ŏ��50%�ɂȂ��������������Ă��邪�A�ǂ̒n����吳�̏����ƂȂ��Ă���B�{��������s�s���̐i�n��Ɠ��l�ȌX���������Ă���A�����̐�ŏ�������A�吳�����ɐ�ŏ��50%�ƂȂ��Ă���B�אڂ��鎭�����́A������旦���Ⴂ���Ƃ��l�����킹��ƁA���B�̓L�c�l�̐�����ɏ\�����ӂ���K�v������B�܂��A�s�s���ȊO�̐�ł̗v���i��A�u�a�A�X�ї��ȊO�̊������j�̋������K�v�ł���B���͓s�s���̐i�n��ł���Ȃ���A�}�Ȃ���������̃O���t�������A���a20�N��ɐ�ŏ��50%�ɂȂ��Ă���B���������Ɠ��l�ȌX���������Ă���B
�H�c�A�a�̎R�A�ΐ�͐��}���ɐ�ŏ�������Ă���B�����̌��ł́A���̓s�s���ȊO�̋}���ȃL�c�l�̐�����̉��ρi���c�����j�A�_��̎g�p�ɂ��v�����l������B
�����ɂ������ŏ�̒l�͂���ȑO�̐�ŌX���f���Ă���ƍl������B���������s�s���̐i�����ł͂��̎����̒l�����n��ɔ�ł������B�L�c�l�͓s�s���̉e�����Ղ��ƌ����Ă������낤�B
�}�Q�|�R�|19�̓^�k�L�ɂ��Ď��������̂ł���B�^�k�L�̃O���t�͂��������肪�}�ŁA��������ɂ������ŏ�͒Ⴂ�B�����A��ʂ͊����X������邭�A��ŏ��50%�ƂȂ����̂����a�����ł������B���A��t�A�ΐ�A�F�{�͐��}���Ȑ�ŌX���������Ă���B�L�c�l�Ɣ�׃^�k�L�̏ꍇ�A�s�s���̐i�s����������̐�łɌ��т����Ƃ͂Ȃ��ƌ����邾�낤�B�t�ɁA�s�s�����i�݈ꂽ���ł̌X�����݂���Ƌ}���ɐ�ł���ƍl������B�����t�͂��̗�ƌ�����ł��낤�B�ΐ�̓L�c�l�Ɠ��l�Ƀ^�k�L����ŋ�旦�������B�����āA��ł͏��a10�`20�N��ɂǂ���̎���}���ɋN���Ă���B���̒n��͐ΐ�ł��X�ы敪�R�ȏオ�������ł�����A�A�����z�Ƃ͈قȂ��������v�����l������B��ł��X�ы敪�R�ȏオ�������ł�����A�A�����z�Ƃ͈قȂ��������v�����l������B
�ΐ�ƂƂ��ɌF�{���^�k�L�̐����Ɋւ��ėv���ӂȌ��ł���B
�A�i�O�}�Ɋւ��Ă͏���s�����Ă��薾��傤�ȌX�������邱�Ƃ��ł��Ȃ������B
�����Ώێ킩��݂��n��̑��l�����݂��i�Ƃ��āA�܂��e�X�̒n��ɂ����鐶���퐔�̑�������̔��f��Ƃ��邱�Ƃ��ł���B�����A�����퐔�������Ƃ������Ƃ́A���̒n��̐����������ꂾ�����l�����Ă��邱�Ƃɂ�邩��ł���B�Ⴆ�A�����̑ΏۂƂ��ꂽ�W��̚M���������݂Ă��A�q�O�}�A�c�L�m���O�}�A�j�z���U���Ƃ�������́A�[���傫�ȐX�т����̐�����ՂƂȂ��Ă���̂ɑ��A�^�k�L�A�L�c�l�A�C�m�V�V�Ƃ�������͕K�������O�҂��v������قǑ�K�͂ȐX�т�K�v�Ƃ��Ȃ����Ƃ͕��z�}�����Ă����炩�ł���B�ނ�́A���Ԃ̋x����ɐB�ꏊ�Ƃ��Đl�ڂ�����邽�߂̑a�т�������Ƒ����̂ł����āA��Ԃ̍̉a���̑��̍s�����͑唼���ނ���X�ъO�ɂ���Ƃ������^�C�v�̓����ł���B
�^�k�L�A�L�c�l�A�A�i�O�}�����������ƐX�тƂ̌��т��́A����܂ł̕��͂Ŗ��炩�Ȃ悤�ɐX�ї��̑����ɔ���������旦����������Ƃ����͂����肵���X���������Ă���B
����A�^�k�L�A�L�c�l�ł́A�X�ї�40�`70%���s�[�N�ɂ��̑O��A�����X�ї�70%�ȏ�Ɠ�10�`40%�Ő���������������X�����݂���i�}�Q�|�R�|17�j�B�����A�L�c�l�A�^�k�L�͕��z�}�̏�ł͐[���X�т���A�a�сA�_�k�n�Ɏ���L���͈͂ɐ������Ă��邱�ƂɂȂ��Ă��邪�A�X�ї��Ƃ̊W�ł݂�ƁA�����̊�Ղ͒����x�̐X�ї������n��ɂ���Ƃ�����B���̂��Ƃ��炵�Ă��X�ї��Ɛ������Ƃ̊ԂɈقȂ������ւ����M���������d�����Đ�����������͑��l���ɕx��ł���Ƃ����悤�B���̂悤�Ȋϓ_����͂W��S���̕��z�}���d�ˍ��킹�A���{���\����M���������̕��z�l���𖾂炩�ɂ��A���̕��z�p�^�[������e�X�̒n�悪���������̒n��������тɕ��z�����v���ɂ��Č����������邱�Ƃ��]�܂����B�������A�L�c�l�A�^�k�L�A�A�i�O�}�́A�s�X�n�A�Ώ��ȂǓ���̒n��������Ėw��ǑS���I�ɋϓ��ɕ��z���Ă���悤�Ɍ����A���z���������ĂłȂ��A����ɃA�i�O�}�ɂ��Ă̓A���P�[�g�����̒i�K�Ń^�k�L�Ƃ̎��ʂ��ǂ̒��x�m������Ă��邩�ɂ��Ă͋^�₪�c��̂ł��̂R��ɂ��ẮA�Ώۂ��珜�O�����B
�]���āA���͂ɍۂ��Ă͕��z���������m�ȃq�O�}�A�c�L�m���O�}�A�T���A�V�J�A�C�m�V�V�̂T���Ώۂɕ��z�}�̏d�ˍ��킹���s���A�n��̑��l�����тɕ��z�����v���̉�͂����݂��B
�i�Q�j�q�O�}�A�c�L�m���O�}�A�j�z���U���A�V�J�A�C�m�V�V�̏d�����z
�L�c�l�A�^�k�L�A�A�i�O�}�̂R��������q�O�}�A�c�L�m���O�}�A�j�z���U���A�V�J�A�C�m�V�V�̂T��̕��z�}���d�ˍ��킹��ƁA�ő�S�킪���ׂďd�����ĕ��z���Ă���n��ƁA�ǂ̎���S�����z���Ă��Ȃ��n�悪���m�Ɏ������B
�k�C���ɂ͏�L�T��̂����q�O�}�ƃV�J�̂Q�킵���������Ă��Ȃ��̂ŁA�ő�Q��̏d�����������݂��Ȃ���ł��邪�A�Q�킪���Ȃ�L��ɂ킽���ďd�����z���Ă���n��́A��ʂ��đ��R�n�̐Ύ�x�A�\���x�̓쑤�j�y�\�c�R�A�E�y�y�T���P�R����\���R���̊�A�돟�����z���ė[���x�A�������A��ʒ��ɂ����Ă̈�сA�\���R�������A����A�V���A�Y�͒��̎R�x�n�сA�\���R�������A���D���A����A�L�����̎R�x�n�сA���ΘH�A���y�A�P�q�{�A�Y�����x�����Ԉ�сA�N�ʁA�����A�k����̏�����ԎR�n�сA�Η��x����m��������ɂ����Ă̈�сA�㒃�H�𒆐S�Ƃ����сA�V���x�A���x�A���ʊx�ȂǂƂȂ��Ă���B���̂悤�ɂ��Ėk�C���ɂ�����Q��̕��z�l�����݂�ƁA��r�I���ʐς̕��z�悪����ɎU�݂��Ă���̂������Ƃ�����B
����A�{�B�ɂ̓^�k�L�A�L�c�l�A�A�i�O�}�̂R��������ƁA�c�L�m���O�}�A�V�J�A�j�z���U���A�C�m�V�V�̂S�킪�������Ă���A����炪���ׂčL���͈͂ŏd�����Ă���n���Ƃ��ẮA�ʐς̍L�����ɎR���A����A�É��̂R���ɂ܂������A���v�X�R�n�̋�P�x�A�����x�A���x�A�喳�ԎR�Ɏ���S��A�O�d�A�ޗǁA�a�̎R�̂R���ɂ܂����鍑���R�A��䂪���A�߉ނ��x�A����q�x�Ȃǂ̎R�x�n�сA���s�{�A����A���ꌧ���̎O���x�A���ނ��x�A��ǎR�𒆐S�Ƃ����~�A�����ŌQ�n�A��ʁA�����s�A����A�R�������̉_��R�A�b���M���x�A�肪�x��т̎R�x�A���h���x���ӁA�_�ސ쌧�O��R���~�A���\�����R�쑤�ƂȂ��Ă���B
�i�R�j�j�z���U���A�V�J�A�c�L�m���O�}�̏d�����z��
���ɁA���z�̒��S�͎R�n�̐X�тł�����̂́A���c�┨���܂ޒ�n�܂ōL�����z���Ă���C�m�V�V���������A����ΐ^���̐X�сi�R�n�j���M�������ł���B�j�z���U���A�V�J�A�c�L�m���O�}�̂R��̏d�����z���E�o�����Ƃ���A��L�S��̏d�����z��ȊO�ɂ́A��茧�̌ܗt�R�A�Ȗ،��̒j�̎R�A�������ӂ��V���ɕt������ꂽ�B
�E�o���ꂽ�j�z���U���A�V�J�A�c�L�m���O�}�̏d�����z��̋�搔�i�Tkm�~�Skm���b�V���j���\�Q�|�R�|18�̂Ƃ���ł���A�k�C������������搔12,367�̂킸��1.8%�@238���ɂ����Ȃ������B
�܂��A���ꂼ��̎�̐�����搔�i�j�z���U���F2130.5�A�V�J�F1963.5�A�c�L�m���O�}763.0�j�ɑ����́A11.2%�A12.1%�A31.2%�ł������B
�d�����z��́A���k�i�ܗt�R�̂݁j�E�֓��E�����E�ߋE�̂S�n���Ɍ����Ă���A��搔���猩��Ɗ֓��E�����E�ߋE�ɕ��Ă���B��B�n���ɊF���ł���̂́A�c�L�m���O�}����ł������Ƃɂ��B�l���E�����n���ł́A�����R��̂��ꂼ��̕��z�悪�����A�d�����z�悪�����Ȃ������B
���k�n�������Ȃ��̂́A�V�J�̕��z�悪�����Ă���i���؎R�|�{��A�ܗt�R�|���j����ł���B
�d�����z�悪�A�C�m�V�V�̎��R���z����i�{���������邱�Ƃ����ԓI�ɉ\�ŁA������������l�ׂ̉e�����Ȃ��Ɖ��肵���n��j�ɂ���A���̏d�����z��ɂ̓C�m�V�V����������Ƃ����W��������B���̊W�́A�^�k�L�E�L�c�l�E�A�i�O�}�ɂ��Ă����l�ł���B
�ȏ�̂��Ƃ���d�����z��́A�䂪���̑�E���^�M���ނɂƂ��Ă̐����v���������Ă���ƍl������B
�d�����z�悪���݂���̂́A�����R�n�A�ԐΎR���A��ˎR�n�A�I�ɎR�n�Ȃǂ̎R�x�n�тł���B
�����n��̐A�����݂�A�㕔�͈����ѐj�t���ёсA�����̓u�i�тő�\�����≷�ї��t�L�t���ётɏ����邪�A�I�ɎR�n�̏d�����z��݂̂́A�����ɍL���g���я�L�t���ёт��܂�ł���B
�܂��A���̈ʒu�́A�Q�`�S�s�{���̌����t�߂ɂ���A�l�̐ڋ߂�j�ނ悤�Ȏ��R���̍����n��ł���B
���������āA����̒��E��^�M�������̐������\�ȑ��l���ɕx�ފ��́A�䂪���ł͂��̂悤�ȎR�x�n�т��܂ޒn��ɂ̂ݎc����Ă���A�Ƃ�����B
�@
�@�{���ł͎�ʂ̕��z�ɂ��ẮA���{�Ƃ����L�����n���I��{�P�ʂƂ��Ď戵���A�n��Ԃ̔�r�͌����Ƃ��Ēn�����x���ōs���Ă������A�����ł́A�����Ώێ�S�̂̕��z��s���{���Ƃ����A�l�Ԑ����Ƃ�薧�ڂ����n��P�ʂŔc�����邱�Ƃ����݂��B
�@�����y�ѐ�ŋ�旦�����A�s���{����ޕʂ��A����Ɋ�Â��s���{�����ƂɃ��[�_�[�`���[�g���쐬�����B
�@����܂ł̕��͂Ŗ��炩�Ȃ悤�ɁA�Ώێ�̕��z�́A�n�j�I�v�����ϓ��̎��R�v���ƂƂ��ɁA�n��̐X�ї������ɑ�\�����l�דI�v���ɂ���ċK�肳�����̂ł���B
�@���������Ė{�`���[�g����͓s���{�����ƂɑΏێ�̎��R���z�̏�A��ς̉e���A�l�דI�e���̓x����ǂ݂Ƃ邱�Ƃ��ł��A����ƂƂ��ɁA����܂ł̕��͂ł͍l������Ă��Ȃ������X�т̎��I����ɂ��āA��̕��z�ɑ��ĈقȂ�e����^���Ă��邩�ǂ����������邱�Ƃ��\�ƂȂ�B
�i�P�j�����Ώێ킲�Ƃ̐����ɂ��s���{���̗ޕ�
�����Ώێ�ʂɐ�����旦�A�ɐB��旦�A��ŋ�旦���ɂ��A�s���{����ޕʂ������ʂł���B�������e����ɂ���ĈقȂ��Ă��邽�߁A�ޕʋ敪���͂X�`�T�܂Ŏ�X�ł��邪�A������������N���i���[�}�����\���j�̏������قǁA������Ԃ͗ǍD�ł��邱�Ƃ������Ă���B
�\�Q�|�R�|19�`26�́A�Ώێ킲�Ƃ̗ޕʌ��ʂŁA�\�Q�|�R�|27�͂�����ꊇ���ĕ\�킵�����̂ł���B
|
�\�Q�|�R�|19�@�j�z���U���̐�����旦�A�Q��^������旦����݂��s���{���̗ޕ� �\�Q�|�R�|20�@�j�z���W�J�̐�����旦�A���N�^������旦����݂��s���{���̗ޕ� �\�Q�|�R�|21�@�c�L�m���O�}�̐�����旦�A��Ł^������旦����݂��s���{���̗ޕ� �\�Q�|�R�|22�@�q�O�}�̐�����旦�A��Ł^������旦����݂��s���{���̗ޕ� �\�Q�|�R�|23�@�C�m�V�V�̐�����旦�A��Ł^������旦����݂��s���{���̗ޕ� �\�Q�|�R�|24�@�L�c�l�̐�����旦�A��Ł^������旦����݂��s���{���̗ޕ� �\�Q�|�R�|25�@�^�k�L�̐�����旦�A��Ł^������旦����݂��s���{���̗ޕ� �\�Q�|�R�|26�@�A�i�O�}�̐�����旦�A��Ł^������旦����݂��s���{���̗ޕ� |
�i�Q�j���E��^�M���ނ̑����I�Ȑ�������݂��s���{���̗ތ^��
�{�����Ŗ��炩�ɂ��ꂽ���E��^�M���ނW��i�����ł̓q�O�}�A�c�L�m���O�}�͒P�ɃN�}�Ƃ��ē����ވ����Ƃ����j�̐�������A�s���{�����^�C�v�������邽�߁A���̎菇�ɂ�背�[�_�[�`���[�g���쐬�����B�i�}�Q�|�R�|20�j�B
�P���_�𒆐S�ɓ��p�x�i360/�V�K�j�Ő��������A�����10�������A�Q���̏�ɕ\�Q�|�R�|27�Ɏ��������N�ɏ]���v���b�g�����B�R���̏ꍇ�A��ɂ�胉���N�����قȂ邽�߁A����Ƀv���b�g����ʒu�͉��}�Ɏ����Ƃ���Ƃ����B
|
�Ȃ��A���Y�s���{���Ɏ��R���z���Ȃ��킪����ꍇ�́A���̎��\�킷�����폜�����B
�C�D���[�_�[�`���[�g�ɂ��s���{���̗ތ^��
���[�_�[�`���[�g�̃p�^�[���ɂ��܂����̃^�C�v�ɗޕʂł���B
�A�D�^�C�v�`�F�����Ώێ�̂��ׂĂ������x���ŕ��z����B
�P�@�`�|�P�@�����x�����a�^
�Q�@�`�|�Q�@��x���@�V
�C�D�^�C�v�a�F�Ώێ�͂��ׂĐ������邪���̕��z�ɕ肪����B
�P�@�a�|�P�@�N�}�����^
�Q�@�a�|�Q�@�N�}�E�V�J�����^
�R�@�a�|�R�@�T���E�V�J�E�C�m�V�V�����^
�S�@�a�|�S�@���̑�
�E�D�^�C�v�b�F���R�I�v���A�l�דI�v���ɂ�舽���̕��z�������Ă�����́B
�P�@�b�|�P�@�N�}�����^
�Q�@�b�|�Q�@�C�m�V�V�����^
�R�@�b�|�R�@�V�J�E�C�m�V�V�����^
�S�@�b�|�S�@�V�J������
�T�@�b�|�T�@���̑�
�������Ƃ�܂Ƃ߂����̂��\�Q�|�R�|28�ł���A����Ɋe�^�C�v�������x�����a�^���o���_�Ƃ��Đ��肳���o�߂�ǂ��Ĕz���̂��}�Q�|�R�|21�ł���B
���̐}�y�ѕ\���疾�炩�Ȃ̂́A�c�L�m���O�}�����ތX�����������A���̕��z�������{��������߂đ������Ƃł���B�����ő����̂̓N�}�E�V�J�̂Q�킪���ތX�����������ŁA�V�J�P�Ƃ̐��ނ��邢�͕��z�̌����͂R���ŔF�߂���ɂ����Ȃ��B���ڂ��ׂ��Ȃ̂́A�j�z���U���ŁA���̐X�ѐ��M���ނɐ�Đ��ތX�������������͑S���Ȃ��A�{�킪���ތX�����������A���z�������ꍇ�́A�V�J�E�N�}�������̓C�m�V�V�i���R���z���Ȃ��ꍇ�������j�������X���������B
�ȏ�̂��Ƃ��璲���Ώێ�W��̂����ł̓c�L�m���O�}���n��I�ɍł������p�������Ղ��A�����ŏ��ł���\���������̂̓V�J�ł���A�T����C�m�V�V�͖{���̕��z�悩��p���������Ƃ͌����_�ł͂���߂ċH�ł���A�Ƃ������Ƃ��ł���B
�s���{���ʂɂ݂�ƁA���E��^�M���ނ��n��I�ɍL�����z���Ă���A���E��^�M�����������ł��L���ł���̂́A����A�R���A�ޗǂ̂R���ł���Ɏ����̂́A�O�d�A�a�̎R�A���s�A����A�̊e�{���ł���B�����̕{���́A�N�}�A�V�J�A�C�m�V�V�̂����ꂩ�̕��z������̂����苷���Ȃ��Ă���B
�Ȃ��A�C�m�V�V�����^�̂R���i���A�R�`�A�V���j�͎��R�����ɂ��C�m�V�V�����z���Ȃ����Ƃ������A���E��^�M���������́A��L�̕{���Ɠ����x�ɖL���ł���Ƃ����悤�B
��ʁA�����A�_�ސ�̓s���́A�V��̚M���ނ���x���ł��邪���ׂĕ��z���Ă���B����͕��n�A�u�˕��͊J�����ꂽ���̂́A�傫�ȎR��ɂȂ���R�n�����݂��Ă��邽�߂Ǝv����B�R�n�����������Ǘ����Ă���A���{�̂悤�ɃN�}�̕��z�����@�����`�ƂȂ�B���茧�͒��E��^�M���ނ̐��������X�ɂ��߂�ꂽ��Ԃ��l������B��錧�̓N�}�A�V�J�A�T���̎R�n���M���ނ̕��z���������`�ƂȂ��Ă���A�n�j�I�Ȑ���̂���k�C���A����������Β��E��^�M���ޑ��̍ł��n��Ȓn��Ƃ�����B
�@1978�N�A��Q�R���ۑS��b�����̈�Ƃ��āA���E��^�M���ނW�킷�Ȃ킿�j�z���U���iMacaca fuscata�j�A�V�J�iCervus nippon�j�A�c�L�m���O�}�iSelenarctos thibetanus�j�A�q�O�}�iUrsus arctos�j�A�C�m�V�V�iSus scrofa�j�A�L�c�l�iVulpes vulpes�j�A�^�k�L�iNyctereutes procyonoides�j����уA�i�O�}�iMeles meles�j�̕��z�������s��ꂽ�B1979�N�ɂ́A�������ꂽ�W��̕��z�Ɋւ�����͎��C�e�[�v�ɓ��͂����Ƌ��ɁA�Tkm���b�V���\���ɂ��S�����z�}���쐬���ꂽ�B1980�N�́A���C�e�[�v�ɓ��͂��ꂽ�����R���s���[�^�|�ɂ���ďW�v�E�������A���̍��y���l���Əd�˂��킹�āA�킲�Ƃ̕��z�X���ׂ��B����ɁA1979�N�ɓ���ꂽ���z�}�Ɛϐ�Ȃǂ̊��v���Ƃ��ׂ��킹�邱�Ƃɂ���āA�e�X�̎�̕��z�����肷��v������͂����B�܂��A�e�X�̎�̕��z�}���d�˂��킹�邱�Ƃɂ��A�쐶�M���ނ̕��z�悩��݂����R�n��̉��l�Ƃ��̓��Ԃɂ��ĕ��͂��s�������ʁA�ȉ��̂��Ƃ����炩�ƂȂ����B
�i�j�z���U���j
�j�z���U���̕��z�ɉe�����y�ڂ��ƍl��������v���͂������ʁA�i�P�j�j�z���U���́A1.5m�ȏ�ϐ�[����50���ȏ�̑���n�тɂ��A���̂���̏��Ȃ��n��Ƃ����Ȃ����z����A�i�Q�j�����їт̑��݂́A�j�z���U���̕��z�𐧌�����A�i�R�j�j�z���U���͐X�ї��̍����n���I�D���邱�Ƃ����炩�ɂȂ����B
�i�V�J�j
�k�C���A�{�B�Ƃ��V�J�̕��z���K�肵�Ă����`�I�ȗv���͐ϐ�[�ł��邪�A�k�C���Ɩ{�B�ł͕��z�𐧌�����ϐ�ʂ͈قȂ��Ă��邱�Ƃ����炩�ƂȂ����B
�z���V���E�W�J�́A50cm�ȏ�ϐ�[��20���ȏ�̒n��ɂ͂قƂ�Ǖ��z���Ȃ��B�G�]�V�J�̕��z�����������̂́A60cm�ȏ�ϐ�[��80���ȏ�̒n��ł���ƍl������B�V�J�̕��z���S�́A��̏��Ȃ��n��ł��邪�A�����̒n��̒��ŃV�J�̕��z�悪�����n��ł́A�V�J���ߑ�ɗl�����ƍl������B�ߑ�Ȏ���ƐX�ђn��̂��܂��܂Ȍ`�ł̊J���́A�V�J�̌̌Q������ɏ��������Ă���B
�i�c�L�m���O�}�j
�c�L�m���O�}�̕��z��̓u�i�E�~�Y�i���N���X��̐A���ƑΉ����Ă���A�c�L�m���O�}�͂��̐A�����傽�鐶���n�Ƃ��Ă���ƍl������B
�i�q�O�}�j
�k�C���ɂ�����q�O�}�̕��z��͂U�n��ɕ����邱�Ƃ��ł���B�����̂Q�n��̌Q�́A���̂S�n�悩�犮�S�ɐ�͂Ȃ���Ă���B�q�O�}�̌��݂̕��z��́A�X�ї�40%�ȏ�̒n��Ɍ����A���z�̒��S��͓����̐X�т�57%���߂鍑�L�тł���B
�i�C�m�V�V�j
�C�m�V�V�̕��z�K���v���́A�V�J�Ɠ��l�ϐ�[�ł���A�{��̕��z��k���̕��z�O���͐ϐ�[30cm�ȏ�̓���70���ȏ�̒n��̋��E���Ƃقڈ�v����B�{��̏o���Ɛ�ŏ��̕��͂̌��ʁA�n�����z�̒��S�́A�{�B�ł͋ߋE�n���A�l���ł͈��Q���A��B�ł͋{�茧�ł���A���k�A�k���A�k��B�n���͕��z���ӂƍl����ꂽ�B���{��̓V�J�ƈقȂ�A�_�k�n�A����A��R�тȂǂɂ��L�����z���Ă��邪�A���̔w��ɂ͕K���R�т����݂���B
�i�L�c�l�A�^�k�L�A�A�i�O�}�j
�R��Ƃ��ɂ��̕��z�́A�X�ї��R�ƂU�i���Ȃ킿10�`70%�j��I�D����X�����������B
��s�s�i�����A���Ȃǁj���ӂł́A�L�c�l�A�^�k�L�Ƃ��X�ї��X�i70%�ȏ�j�̒n���
�I�D����A�Ƃ����I�ȑI���X�����������B
�@�L�c�l�A�^�k�L�A�A�i�O�}�̐�ŏ��̔N��I���ڂ�����ƁA�R�틤�ɓs�s���̉e����
�����Ă��邪�A���ƂɃL�c�l�͓s�s���̉e���������₷���ƌ����悤�B
�i�d�����z��j
�@�k�C���ɂ�����V�J�A�q�O�}�̂Q��A�{�B�ɂ�����j�z���U���A�V�J�A�c�L�m���O�}�A�C�m�V�V�̂S��̏d�����z��́A��������A�[���R�x�n�т����S�ƂȂ��Ă���B
�i�s���{���ʂ̐����j
�@�Ώێ�S�̂̓s���{�����Ƃ̐������A���[�_�[�`���[�g��p���đ����I�ɔ�r���������Ƃ���A���E��^�M�����������L���Ȓn��ƕn��Ȓn�悪���Ăɋ�ʂ��ꂽ�B�L���ȓs�{���͂�������R�A�S��̏d�����z������n��ł������B
���s���{���Ԃɂ݂�ꂽ��X�̐����p�^�[���́A�J���̐i�W�ɔ����A�M�������̐�����̏k���̎��n��ω���\���Ă���ƍl����ꂽ�B
�i���������j
�@�䂪���̒��E��^�M���ނ̂����A�����ς�X�тɈˑ������̌Q�̈ێ��ɂ͍L���n���K�v�Ƃ���j�z���U���A�V�J�A�c�L�m���O�}�A�q�O�}�́A�e��̊J���s�ׂɂ�萶���悪���߂���ƂƂ��ɁA�c���ꂽ��������Ǘ�������X���ɂ���B�Ǘ���������������̌̌Q�͑傫�Ȋ��̕ω��i��A����j�ɂ���Đ�ł��Ղ��A���̂悤�ɂ��Đ������n��͎��ӂ̌̌Q�ɂ���Ė��߂邱�Ƃ�����Ȃ��߁A��Œn��̊g�傪�i�݂₷���B
�쐶�����̕ی�E�Ǘ��ォ��́A�����n�̌Ǘ�����h���z�����K�v�ł��낤�B
�l�Ԃ̐������̎��ӂ��邢�͓����ł̐������\�ȃC�m�V�V�A�L�c�l�A�^�k�L�A�A�i�O�}�Ȃǂ̐����͑O�L�S��ɔ�ׂ�A���肵�Ă���Ƃ����Ă悢���A�s�s�����i�n��ł́A���̐�����������Ȃ��Ȃ��Ă���B
�@