IV. 総括と展望
IV. 総括と展望
今回の調査結果のとりまとめについては、干潟、藻場、サンゴ群集のそれぞれについて個別に考察が行われている。本稿では、結果の総括の性格上統計について多少の重複を含みまとめると共に、一斉調査結果だけではうかがえぬ各棲み場所類型の減少、荒廃の原因についてわが国における個別研究の結果を引用して考察を試みた。また、一斉調査における手法の問題点、今後の展望について議論した。
1. 干潟
(1)干潟の現存面積
今回の全国一斉調査で把握された現存干潟の総面積は51,443haであった。海域的には大規模な閉鎖的内湾で多くの河川が流入する有明海が最も広大な干潟を有しており、有明海全体の干潟面積は20,713haに達し全国の干潟面積の約40%を占める。単一の干潟としては熊本県有明海に位置する前浜干潟で1,656haの砂質干潟である。有明海奥の福岡県〜佐賀県地先には軟泥質の1,000haを越える干潟があり、有明海内の枝湾である長崎県の諌早湾にも1,846haの泥質干潟がある。有明海に次いで現存干潟面積が大きいのは瀬戸内海西部の周防灘西海域で6,409ha、以下八代海の4,465ha、東京湾の1,640ha、三河湾の1,549ha、伊勢湾の1,395haの順であり、そのほか海域別で合計面積が1,000haをこえる干潟が存在したのは、沖縄の1,216ha、北海道の根室地区1,148ha、網走地区1,091ha、瀬戸内海燵灘の1,022haで、以上の10海域で日本全体の干潟の約80%を占める。
(2)干潟の消滅状況と原因
前回(1978年)調査後約13年間に消滅した干潟の総面積は3,857haであり、大規模なものは有明海、別府湾、東京湾、伊勢湾、沖縄島、八代海で認められた。海域別には現存干潟面積最大の有明海が喪失面積でももっとも大きく、1,357haと全消滅面積の約35%を占めた。次いで別府湾と東京湾のそれぞれ281ha、280haが大きい。消滅原因のほとんどは人間の開発に起因するものである。有明海の消滅域のうち約86%は陥没によるものとされ、単一の干潟として最大の陥没喪失は福岡県柳川市昭代地先の前浜452haである。今回の調査では陥没の原因については言及していないが、この海域は、三池炭田の海底炭田開発が多年にわたって行われており、干潟面の広域陥没はこれとの関連を予測させる。それ以外の海域では主な消滅理由は埋め立てでそれに浚渫が複合している。
理由は空港建設、都市や港湾の再開発、発電所立地造成などさまざまである。兵庫県では、消滅理由でその他として挙げられているものが46haで83.6%を占めているが、砂の自然流失によるという。これだけでは判断し難いが、天然の干潟や砂浜の砂が近年急に流失することは考え難く、近接域の地形変更による流況の変化や海底の砂採取などが残った干潟の砂の流失に影響した可能性を検討する必要がある。わが国の農業環境の変化に伴い干潟消滅の理由としての干拓はほとんど影をひそめ今回の調査では僅かに三重県で84haの1件をみるのみである。しかし、長崎県の諌早湾では、最後の大型農業干拓として、沖合い締切堤で3,550haを囲い込み1,846haの泥質干潟のうち1,550haを干拓地及び調整池に変える計画が、平成元年度以後着工され現在進行中である。
2. 藻 場
本調査で藻場としてとりあげた海産植物群落は、その構成種からみてアマモ場(水中顕花植物であるアマモ属、リュウキュウスガモ属、リュウキュウアマモ属、ボウバアマモ属など)、ガラモ場(ホンダワラ科大型褐藻)、コンブ場、アラメ場(アラメ、カジメ、クロメなどの大型褐藻)ワカメ場、テングサ場、アオサ・アオノリ場の7タイプからなっている。沿岸の諸生物の構造的生息場所型として広域かつ永続的で重要なのはアマモ場、ガラモ場、コンブ場とアラメ場を併せたいわゆるケルプ群落である。これらのうち海草のアマモ類だけが砂泥底に生え根を通じて海底から栄養塩類を吸収する。それ以外は主に岩礁、転石といった硬基質上に生育する。テングサについてはガラモ場、コンブ場、アラメ場、ワカメ場などの大型藻類の下生えとして、岩礁上面に丈低く密生するもので、その生育の良否は前述の大型藻類の消長とむしろ相反関係にある。アオサ・アオノリはほかの藻場構成種とは性格を異にし、本来群落遷移では初期段階に出現し長期永続性を欠くとされているが、高窒素環境を好み、近年富栄養海域でしばしばアナアオサが大規模に繁茂する事例が多くなっている。このアナアオサは硬い基質上に付着して生育するだけでなく、脱落後もかなり長期間沿岸に堆積あるいは浮遊しながら生きており、次第に枯死しながら海底面を覆って海中生物の生活に負の効果を及ぼすことが指摘されている。
(1)藻場の現存面積と消滅状況
本調査の全国藻場総面積は201,212haとなっており、前回(1978年)調査後の約13年間に6,403ha(現存藻場面積の3.2%)が消滅している。全国を91海区に区分して海域別の変化を調べたところでは、消滅藻場面積が最大の海域は天草灘(九州西岸)で全国の14.8%を占めており、第2位以下は秋田の11.8%、日向灘(宮崎県)の6.8%、有明海(九州西岸)の6.7%、播磨灘北(瀬戸内海)の5.9%、と続いている。瀬戸内海は多くの海域に分けられているが紀伊水道、豊後水道を含めた全瀬戸内海域の消滅藻場面積は全国の20.8%に及び依然としてこの海域での減少傾向が鈍っていないことを示している。
消滅藻場面積をタイプ別に集計すると全国の消滅藻場面積の22.2%をガラモ場が占め、次いで19.9%をアマモ場が、16.2%をアラメ場が14.9%をワカメ場が、13.3%をテングサ場が占めているが、個々の海域別の消滅藻場のタイプ別構成はかなり異なっており、海域別の性格、原因を考えて行かなければならない。九州西岸、同じく東岸の日向灘などでアマモ場の減少が著しいが、この海域は生物地理的にみてアマモZostera marina の分布南限に位置しており、埋め立て、汚濁など人為改変によらない死滅、藻場面積の減少については、実態はあいまいながら海洋気候の長期変動と植物側の活性の変化を考慮する必要があるのかも知れない。このことについては、後段で触れたい。
(2)藻場の消滅状況と原因
現存藻場面積に対する消滅藻場面積の割合すなわち消滅比率を集計した結果ではその大きい方から1位、2位、3位、4位及び6位が瀬戸内海沿岸にあり、藻場衰退がこの水域の自然環境劣化の大きな課題であることを示している。第7位は三河湾(愛知県)であるがこれも沿岸地域の都市化が進み富栄養化が進んだ地域である。日向灘(宮崎県)、有明海(九州西岸)、鹿児島湾(鹿児島県)と沿岸の人為開発が比較的小さい南西九州で消滅比率でいずれも10位以内にランクされているのは今回の特色といえる。実面積においてもこれらの諸地方の大部分の藻場喪失面積は数百〜数千haに及んでおり、減少が著しかった事になるが、一方現存藻場の面積でみると、瀬戸内海、西南九州は依然として全国的にみて高い順位にある。以前から藻場面積が大きく今回の調査でも減少が少ないのは熊野灘、(アラメ場、テングサ場、ガラモ場)、駿河湾(ガラモ場、アラメ場)、能登半島(ガラモ場、アマモ場)、北海道沿岸の諸地方(アマモ場、コンブ場)などである。
藻場消滅の原因としては、全国的には、埋立等直接改変が28.1%とされているが、全国の消滅藻場面積の約21%を占める瀬戸内海とその周辺域ではその比率が43%に達している。また面積としての絶対値はそれほど大きくないものの埋立等直接改変が消滅原因の100%を占める海域が91海域中26(28.6%)を数える。
一方、藻場の解析を担当した横浜、相生両氏は各県の報告中大型海藻群落の消滅理由として挙げられた磯焼け、及び海況変化が現段階では科学的根拠薄弱として原因不明に含めて考えているが、これらが原因の71.5%に達することは重大であり、今後の藻場の動向を予測する上にも科学的因果関係の解明への努力が必要である。
磯焼けは潮下帯岩礁の大型海藻が海流変化に伴う水温の上昇や時化による激しい攪乱、淡水流入などによって死滅したのち、無節石灰藻類が繁茂してしまい(coralline flatサンゴモ平原の形成)大型藻が生えなくなる現象である。元は南伊豆地方の漁民の間で用いられた漁業上の用語であるが北日本で水産上問題になっている磯焼けも生物学的にほぼ同じで、石灰藻による被覆が大型海藻群落の回復を阻み長期的に石灰藻群落が持続する現象は世界各地で共通してみられるという認識がひろまってきた。いったん無節石灰藻の被覆ができてしまうと、その状態は物理的条件が回復しても数年、または十数年にわたって持続する例が知られている。近年東北水産研究所の藻類増殖室谷口和也室長、および共同して北日本の磯焼けを研究している水産研究所、水産試験場の研究者達は、磯焼けの過程を大型藻類の喪失をもたらす第1段階と、その後無節石灰藻の占拠が長期持続する第2段階に分け、後者の機構として、石灰藻の存在が大型藻の胞子の着底成育を阻害するほか、石灰藻が分泌する化学物質がウニ、藻食性腹足類などの大型グレイザーの稚仔の着底を促し、これらのグレイザー、特にオオバフンウニ属のウニの存在がコンブなどの発芽直後の幼植物を絶えず除去する結果、無節石灰藻に覆われた磯焼けが持続するとみている。そこでコンブ場を回復させる管理技術としてグレイザーを制御することがコンブ場の復元に有効であるとしてグレイザーの食害を生物的にコントロールする手法を開発中である(谷口・関・蔵田、印刷中)。しかし、これも局地的環境改善技術として水産的に有用ではあっても、大規模に磯焼け現象が起こることは防止困難であろう。谷口(1994)は大型海藻の消滅のあと石灰藻の繁茂が起こる場合のほか、汚水排出、陸上からのシルト、粘土などの流入など人間活動の影響としての水質悪化が消滅のきっかけとなり、その場合にはサンゴ藻平原さえも形成されない事に触れ、これに「磯荒れ」という呼び名を与え「磯焼け」と区別することを提唱している。
(3)海水温の長期変動と大型海藻(海草)の消長
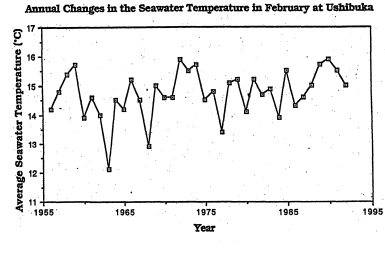
ケルプ類やホンダワラ類といった大型海藻群落は海水温が高い年には成育が悪いことは経験的にかなり以前から知られており「磯焼け」の古典的例とされる伊豆半島の場合は黒潮の大蛇行による異常高温年に大型藻の消失、石灰藻の繁茂が起きるとされている。暖流軸の変動とは別に沿岸の海水温の長期変動で温暖化が進めば、水温変化と結び付いた海藻場の面積減少も生じ得る。図8は熊本県天草の牛深沿岸における冬季二月の平均水温の40年間の変動を野島(1993)がまとめたものであるが、数年に一度の寒冷年はあるものの全体としてのこの十数年は温暖な年が続きやや昇温傾向がみられる。天草灘はこの13年間にガラモ場の面積減少が著しいことが指摘される海域であるが、その主な海域は牛深市西側にあり、野島の観察では以前ガラモ場であった岩礁底にサンゴ群集が広がり若いサンゴ群体が多く成育してガラモを置き換えた状態になっているという。サンゴ礁非形成域では太陽光がとどく浅い岩礁底はサンゴと大型藻類の空間をめぐる競争が激しく、温暖年の持続は造礁サンゴに有利に、寒冷年の持続は海藻群落に有利に影響するものと考えられる。天草灘沿岸は元からアマモ場の少ない海域であったが、天草北西部のアマモ場は1970年代前半に特にめだった人為影響なしに激減し(菊池、個人的観察)、この13年の間に有明海、八代海のアマモ場が激減したのはこの地方がアマモの地球規模の分布の南限域にあたることと合わせ考えると温暖年の継続が衰退の一因になっているかも知れない。ただしこのような水温の経年変動をただちに現在全地球気候の温暖化傾向と結び付けることは差し控えたい。アマモへの高水温の悪影響を論じるためには高水温期の永年変動の情報が必要であろう。
3. 造礁サンゴ群集
造礁サンゴの分布に関し日本の沿岸は、冬季水温が20℃を下らずサンゴ礁を形成する琉球列島(沖縄県と鹿児島県のトカラ列島、奄美諸島)、小笠原群島(東京都)と、サンゴ礁はできないが造礁サンゴ群集は形成する屋久島・種子島以北の暖流影響域(以下サンゴ礁非形成域と呼ぶ)に大別される。
A. サンゴ礁域
(1)サンゴ礁池内のサンゴの状況
今回の調査では、トカラ列島小宝島以南のサンゴ礁域で礁池内総面積約96,000haを調査し、サンゴ群集の面積、約34,190haという値を得ている。海域別では八重山諸島海域が最大で約19,230haで調査面積の約49%を占めていた。次いで沖縄島海域の約7,050haであった。奄美諸島海域は調査域のうちサンゴ群集の生育に不適な沈水裸岩の面積が50%以上を占め、サンゴ群集の面積は相対的に小さかった。被度階級別にみると八重山諸島海域でも5%以下の+クラスが65%近くに達し、被度50%以上のIII、IVの合計は1630.7ha(8.5%)であった。沖縄島海域の場合は約72%が被度5%以下の低被度であり高被度域は周辺の離島部に限られていた。奄美諸島海域はサンゴ群集の面積は相対的に小さいが、サンゴ分布域では被度階級I(5%以上)以上の比率が63%と沖縄島海域よりかえって高い。
第2回調査(1978年)後、主に埋め立て等により、1502.7haのサンゴ礁が消滅した。沖縄島海域での消滅が最大で1224.0haに達している。
沖縄島のサンゴ礁は1970年代にオニヒトデにより大きな被害を受け、回復過程にも陸上の開発に伴う赤土の流入を始め人為汚染の影響も加わって減少の一途をたどっている。八重山諸島では景観的には白保がもっとも優れ、川平にも高被度の群集がみられるが、日本復帰当時の景観に比べると著しく衰退している。石垣島と西表島の間に発達する石西礁湖は13,000haのサンゴ群集面積を擁するが、面積比率の53.7%は被度5%以下であり、被度階級I、II(5%〜50%)の面積比率は36.4%被度階級III、IVの面積比率は9.9%で、高密度の枝状ミドリイシ類の群集の面積は962haであった。石西礁湖では、1974年ごろから鳩間島周辺でオニヒトデの増殖がみられ駆除努力がなされていたが、前回調査(1978年)以降1980年から1981年にかけて石西礁湖全域にわたってオニヒトデの大発生がみられ(1981年の駆除数520,000を極大として80年から84年まで毎年100,000ないし200,000を駆除)この海域のサンゴの多くが失われた(環境庁、1981、福田・宮脇、1982、松下・御前、1983、宇井、1985、亀崎・野村・宇井、1987、岩瀬・野村、1988)。前述の枝状ミドリイシの大群落はその折に食害を免れたもので近年次第に分布を拡大しつつある。
最新の報告(森、印刷中)によれば、今回の一斉調査以後の1992年以降サンゴの被度は急速に回復しつつあり、1993年には被度階級III(50%)以上の場所の面積比率が30%を越え、'94年にはさらに被度階級III以上の面積比率が40%にまで回復した。現在はオニヒトデもほとんど見られなくなり順調に回復しつつあるようであるが、種組成がオニヒトデ大発生前と同様になるかどうかはまだ分からない。
(2)礁縁サンゴの状況
調査員曳航調査法(通称マンタ法)により礁縁の連続目視調査を実施したところ、各地で礁池よりもより高密度のサンゴが存在することを確認しているが、沖縄本島では被度5%以下の地域が2/3を占め、比較的被度が高い最北端の辺土岬周辺、本部半島北側ではオニヒトデの密度も高く、はっきりした食害痕がみられている。沖縄本島周辺の離島、慶良間諸島、伊平屋島などではサンゴ類の被度は高く、良好なサンゴ群落がみられたが、オニヒトデもこれらの周辺諸島でかなりの密度で発見されており、今後の食害拡大が懸念される。
B. サンゴ礁非形成域
サンゴ礁非形成域でも造礁サンゴ群集の存在が確認された。今度の第4回調査においてマンタ法による調査が行われたのは、鹿児島、宮崎、大分、熊本、長崎、島根、高知、愛媛、徳島、和歌山、三重、静岡、神奈川、東京(小笠原を除く)、千葉の15都県である。内田による解析結果にみられるように被度I(5〜25%)以下の低密度地域は輪郭のとりかたで過大に見積る可能性も高いのでこれを除外すると、被度II(25〜50%)以上のサンゴ群集の面積が30haをこえるのは鹿児島、宮崎、大分、熊本、高知、愛媛、和歌山の7県である。県によって必ずしも全県下をカバーしていない県もありその点を考慮すると、被度II以上の地域が70haを越えるのは宮崎、大分、熊本、愛媛であり、和歌山と鹿児島がこれに次ぐもののようである。被度III(50%以上)が50haを越える大面積の高被度サンゴ群集は大分県(豊後水道蒲江付近)、愛媛県(南宇和海)、和歌山県(紀伊半島南部)で報告されている。
全国的にみると、成育型でもっとも優占するのは卓状(51.7%)のミドリイシ(Acropora)属で外洋に面した南日本ではクシハダミドリイシA.hyacinthus が優占し、やや内湾的環境になるとエンタクミドリイシA.solitaryensis がこれに代わって優占する。伊豆半島になると卓状サンゴは希になり、樹枝状のエダミドリイシA.tumida が主要な地位を占める。卓状型についで優占するのは被覆状で、暖海では塊状をなす種も北限近くや深いところでは被覆状に変化するらしく構成種はさまざまである。
前回(第2回)一斉調査の折にはサンゴ礁非形成域の充分な広域調査が行われていないため約14年間の増減を論じるのは難しいが、今回の調査で知られたサンゴ群集の消滅面積は埋め立て等の改変によるもの約3ha、その他の原因によるもの(不明も含む)約11.7haであり、面積的には広くない。
(1)冬季海水温の経年変動と造礁サンゴの寒冷耐性
サンゴ礁非形成域においては冬季海水温の年変動により寒冷年にはサンゴの大量死滅が起きることが知られている。1963年冬は記録的な寒冷年であり(図12参照)、紀伊半島南部、南四国、西南九州など日本の暖流影響域の広範囲で造礁サンゴの死滅がおこっている。前回の調査はそれからの回復過程にあったと推測されている。前回調査(1978年)から今回調査までの間の異常寒冷年の記録をみると、南紀串本の海中公園周辺において1984年2月から3月にかけ海水温が14℃以下に低下する日が3週間以上続き、それに伴い造礁サンゴの群体が多数死滅したことが記録されている(御前、1985)。調査地の岸寄りゾーンのサンゴ類被度第1位の卓状サンゴであるクシハダミドリイシがもっとも水温低下に鋭敏で、寒冷死群体の大部分をこの種とそれに近縁のオヤユビミドリイシが占め、ハナヤサイサンゴも死亡率が高かったがミドリイシ属でも枝状種やキクメイシ類、キクカサンゴ、イボサンゴなどの塊状、被覆状サンゴにはほとんど被害がなかったという。このように造礁サンゴ分布の北限であるわが国のサンゴ礁非形成域では、数年あるいは数十年に1回起きる異常に寒冷な冬の影響を無視できない。十年毎に1回のサンゴ群集の全国一斉調査ではその間のある1年に生じるこのような自然死亡の要因を発見することは不可能である。一方サンゴ礁域ばかりでなくサンゴ礁非形成域においても、夏季水温が28℃を越えるとサンゴ体内に共生する褐虫藻が脱出してサンゴが白化し死亡することが報じられている。
(2)台風等による大規模自然攪乱による死滅
サンゴが成育する暖流影響域はまた台風が毎年通過する地方でもあり、強大な台風の通過は激しい風浪の作用で沿岸浅所のサンゴ群体を破壊する。野島(1993)は熊本県天草南部のサンゴ群集において、1989年11号台風、1991年の17号、19号台風がエンタクミドリイシの卓状群体の多くを破壊したことを記録している。ただし、水温異常低下や台風時の波浪によりもっとも影響を受け易いこれら卓状ミドリイシ類はほかの造礁サンゴ類にくらべて成長も速く、かつ多くの新群体の加入もあるようで、度々の一時的打撃にも拘らず各地でもっとも高い被度を占めている。
(3)サンゴ捕食動物の食害
高知県における局地的消滅はオニヒトデの食害によるものであり、宮崎県における5haの消滅にはヒメシロレイシガイダマシDrupella flagum の食害が原因の一部とされている。この一斉調査以後1991年から1992年にかけ愛媛県宇和海の海中公園地区においてヒメシロレイシガイダマシが大発生してサンゴを食害し駆除が行われた(須賀、1994)。また熊本県天草南部でも1991年から1992年にかけてトゲレイシダマシMorula spinosa が増殖しエンタクミドリイシに対する食害がめだち、地元ダイバーを組織して駆除が行われた(野島、1993)。
(4)人為影響
人間の開発行為によってサンゴ成育域が明らかに破壊された例は、鹿児島県における埋め立てによる3haの消滅がある程度であり、現存被度5%以下の低密度生息域がかつてはもっとサンゴが繁茂していたのか、その衰退に人間活動が関与していたのかどうかについては今回の資料では検討不能であった。
図12 牛深における最寒月(2月)の表層海水温の40年間の年次変動
(長崎海洋気象台:西日本海況旬報より)。
4. 今回調査における問題点と今後の展望
(1)前回(1978年)調査と今回(1989〜1992)調査の間に干潟、藻場、サンゴ礁ともかなりの面積減少があり、干潟、サンゴ礁の減少のほとんどと、藻場減少の数十%は埋め立てなど直接的な人為的破壊、環境改変によるものであった。サンゴ礁のサンゴ群集の低被度化については1980年代のオニヒトデによる食害の影響が大きいが、沖縄島本島におけるサンゴ群集回復がおそく現在なお全体的に低被度である件については、陸上の開発による悪影響が主な要因だと推測された。今後の沿岸域自然環境の保全については埋め立てなどの開発行為の禁止、抑制など、陸上からの汚濁物質、泥土などの流入を減らす施策を強化する必要がある。
(2)被度階級の判定と面積の推定については、干出時に直接その面積が一望できる干潟がもっとも推定が容易で、最大干潮時の航空写真があれば正確な面積推定も可能である。干潮時干出しない藻場、サンゴ群集については、まず被度区分の判定に主観が入ることは避け難いことであり、さらに全域を一望することができないという制約のため、被度階級+が広くつながって存在するのか、非生息地の岩礁底にとびとびに被度階級+やI、IIのサンゴ群集があるとみなすかは調査者のその時の判断によることになる。調査者の経験、判断基準は共同講習をすればある程度は客観的に近付けることができるとしても、全国規模で多くの団体、調査者が携わる場合避けることのできぬ問題である。また前回調査時との比較の場合、前回の調査が過大であると消滅面積、消滅率もまた大きく見積られることになる。
(3)サンゴ礁域、サンゴ礁非形成域のサンゴ群集とも今回の調査によって、定性的、半定量的情報が集まったことは貴重であり、評価できる。マンタ法による海岸線の連続観察は点の調査から線、面の調査へとカバーする範囲を拡大したという点では有益であるが、調査水域の透明度、調査者の調査能力、生物識別能力、目撃したことを記録する手法、適切な調査線の設定と曳航ボートの運航技術等各都府県でばらつきがあることは当然で、今後、どのように調査の精度を上げ水域毎のばらつきを小さくしていくかが問題である。造礁サンゴの分類同定についてはオーストラリア国立海洋研究所のヴェロン博士の来日調査により日本の造礁サンゴ相の概要がまとめられ、西平・ヴェロン著の日本の造礁サンゴの図鑑も近々刊行されるので、もちろんある程度のトレイニングは必要だが今後の調査能力の向上を期待できよう。
(4)消滅原因については、埋め立て、浚渫といった明らかな人為的原因以外はほとんど確認しがたい。サンゴ礁域において淡水流入や陸地からの赤土の流入がサンゴの成育を抑制することは多くの事例報告があるが、因果関係は未だ科学的に解明されていない。埋め立てのようなあからさまな消滅原因以外の説明については、人がどう受け留めているかではなく、正しい因果関係を求めようとすれば、今回の聞き取り調査の様な形ではない研究、あるいは消滅、衰退を生じ得る環境要因の経年変化の記録、因果関係の解析か必要である。海水温が平年に比して異常に高い夏、異常に低い冬、長期的昇温または降温傾向を海洋気象台、測候所、水産試験場などの定点継続資料から抜き出す作業、豊水年、渇水年、あるいは極端に淡水流入、土壌流出が起きた年、季節と藻場、サンゴ礁の変化の関連を一部地域についてでも把握することは重要である。アマモ場の衰退が富栄養化や透明度、濁度の長期変化と関連するかどうかについても限定された海域での長期モニタリングの資料なしには判断できない。一時的一斉の保全状態実態調査のほかに因果関係を解明する別の側面からの研究プロジェクトを実施し、消滅、衰退原因に対する対策を検討する必要がある。
(5)生物的要因、空間占拠をめぐる競争、捕食者の食害による衰退などについては個々の事例について環境庁の駆除対策事業や関係者による被害状況の経時的調査がおこなわれていることは評価に値する。しかし保全に有害な生物の生態、生活史に関する知見、大増殖のきっかけとなる要因についての基礎研究はまだ著しく弱体で今後の研究振興が必要である。
文 献
福田照雄・宮脇逸朗.1982.八重山群島石西礁湖海域におけるオニヒトデの異常発生について,海中公園情報 56:10-13.
岩瀬文人・野村恵一.1988.石西礁湖域におけるイシサンゴ類、ソフトコーラル類及びオニヒトデ,海中公園情報 80:8-12.
亀崎直樹・野村恵一・宇井晋介.1987.石西礁湖海域のイシサンゴ類及びオニヒトデの動態(1983-'86年),海中公園情報 74:12-17.
環境庁. 1981.浅海における海中景観の保全と活用の推進に関する調査報告書(西表国立公園石西礁湖の保全と利用),95pp.
松下 甲・御前 洋.1983.石西礁湖におけるオニヒトデの分布について,海中公園情報 59:14-16.
森 美枝. 印刷中.石西礁湖におけるイシサンゴ類とオニヒトデの推移,海中公園情報 107:10-15.
西平守孝・J.E.N.VERON.印刷中.日本の造礁サンゴ,海遊社出版.
野島 哲. 1993.造礁サンゴの分布周辺域におけるサンゴ群集の維持機構について,平成2〜4年度科学研究費科学研究(一般研究 C)研究成果報告書 28pp.
御前 洋. 1985.寒波によるイシサンゴ類の生存限界について,-1984年冬の串本海中公園の状況からの解析,海中公園情報 68:15-19.
須賀秀夫. 1994,宇和海におけるサンゴ食巻貝について,海中公園情報 103:9-13.
谷口和也. 1994.磯焼けとは何か? 海藻生態学の立場から,北大大学院環境科学研究科・株式会社エコニクス共催、地球環境シンポジウム“磯焼け”の現況と課題、講演要旨集:16-19.
谷口和也・関 哲夫・蔵田一哉.印刷中.磯焼けの機構と克服技術としての海中造林,原稿26枚、図6,表2.
宇井晋介. 1985.八重山群島石西礁湖海域におけるオニヒトデ、石サンゴ類の分布の推移と現状,海中公園情報 64:13-17.