4. ウサギ目・ネズミ目(兎目・齧歯目)
はじめに
昭和48年度に開始された第1回自然環境保全基礎調査では、動物分布につい
てはわずかに「すぐれた自然」の調査中で、文献等により主要な生息地等につい
て調査が行われたに過ぎない。昭和53年度の第2回調査では一応主要な動物に
ついての分布調査を実施したが、哺乳動物については大型8種に限定して行われ
た。また、昭和59年度の第3回調査では、全哺乳動物を対象として専門家によ
る分布調査が行われた。さらに、平成2年度〜3年度の第4回調査では、専門家
に加え各都道府県の鳥獣保護員、猟友会員の協力による調査が行われた。
このように概ね5年を周期として調査を実施する背景には、各動物の分布が四
囲の環境変化に伴ってどのように変化していくかを探ることにある。しかし、筆
者が担当したウサギ目と齧歯目に属する全動物のうち、数種類は過去の調査で報
告されないものもあり、経年的な分布域の変動については比較できなかったもの
もある。そこで狩猟動物については、環境庁発行の鳥獣統計を中心に、その他の
動物については関連文献をそれぞれ参考にしながら考察を行った。
考察の視点は、帰化種については分布の現状と、これと生態的地位を同じくす
るものとの競合、捕食/被食関係について、在来種については環境との関連から
分布域の拡大、縮小に注目してそれぞれ考察を行った。考察は紙面の都合上、特
筆すべきものに重点を置いた。なお、本文中、特に断わりのないメッシュ数は、
3次メッシュ数のことである。
ウサギ目、齧歯目は全体として情報量が急増した。それはこの自然環境保全基
礎調査が広く国民の間で認識され、理解を深めた結果の産物であると考えられる。
情報量が著しく増えた種は、たとえば、ニホンリス、エゾリス、シマリス、タイ
ワンリス、ナキウサギといった昼行性動物や一般によく親しまれている動物で目
立った。夜行性動物でもムササビ、ノウサギといった知名度の高いものにもこの
傾向が見られ、鳥獣保護員及び猟友会員の幅広い参加を明確に裏付ける結果とな
った。これに対し、専門家でなければ普段の生活の中で名前すら触れることのな
いノネズミ類、モグラ類の情報は、一般的傾向として初回から大きな情報増は見
られなかった。
今回の分布情報の分析を行った結果の反省としては、ノウサギとユキウサギの
情報が北海道と本州で混同されたことである。今後、調査に先立ち「本州にはノ
ウサギが、北海道にはユキウサギが生息することを明確に表示しておくか、ノウ
サギ一本に統一して調査し、後刻、集計の段階で調整するかの選択が迫られてい
る。
次に、本調査の結果として浮き彫りにされた問題点は、タイワンリスの分布情
報が全国レベルで急増したことである。帰化種であるタイワンリスが余りにも急
速に増加しているが、調査者がニホンリスとタイワンリスを明確に識別している
か否かが大変気に懸かることである。当該地域の専門家によるチェックが必要で
ある。この急速な分布拡大が事実とすれば、動物地理学上由々しいことになり、
何らかの対策が急がれる。これと同じ現象がシマリスとエゾリスで見られ、これ
も全国レベルで急増している。本州に限っていえば、ニホンリスとタイワンリス
のように、直接競合する種はいないが、むやみに外来種を放獣したり、ペットを
篭抜けさせたりする背景が問題である。北海道の場合、シマリスと記載があれば
エゾシマリスと分類され、処理されているが、都市近郊の個体については、外来
種であるチョウセンシマリスかどうかを確認する必要がある。殊に、山岳地帯に
住むエゾシマリスとの交雑の危険性があり注意を要する。
これらの事実から動物のむやみな移動を禁止する法的措置もさることながら、
国民の理解を深めるためのキャンペーンが必要であろう。
4-1 ウサギ目
(1) ナキウサギ科
- (1) ナキウサギ : A 分布パターンを表わしている。
本種は北海道にのみ生息する種で、過去の文献によれば、大雪山系の黒岳、
旭岳、忠別岳、十勝岳、富良野岳などの高山帯と、それらの周辺の置戸、然
別湖付近、夕張山系の夕張岳、芦別岳、日高山系の幌尻岳、札内岳などに分
布している(芳賀、1957、内田、1960)。第3回の基礎調査では、22メッシ
ュで分布情報があったが、第4回では26メッシュに増加している。増加箇所
は十勝管内のもので、相変わらず空知管内の夕張岳周辺からの報告はない。
芳賀(1957)は、ナキウサギの分布は一般的には原産地である置戸をはじ
め、然別湖、大雪山一帯、夕張岳、芦別岳、日高山脈その他といわれている
が、夕張岳や芦別岳ではもうほとんど姿を見ることができないと述ベている。
このことから、夕張山系では絶滅したとも考えられるが、本調査では分布し
ていないことと、情報が得られなかったこととの区別が明確でないところか
ら、実態をさらに混沌とさせている。この点を明確にするための追跡調査が
必要である。
(2) ウサギ科
- (1) アマミノクロウサギ : B やや情報不足。
本種は1属、1種からなり、奄美大島と徳之島にのみ分布している。筆者
らが1972年に両島を調査したとき、奄美大島の湯湾岳、ヤクガチョボシ岳な
どを中心に、徳之島では剥岳、井之川岳、大田布岳などを中心に、それぞれ
広い範囲にわたって生息していることが確認されている。
桐野(1977)によると、標高695mの湯湾岳と、646mの井之川岳の山頂
部は、3000〜4000年ほど前に、日本列島の一部分が水没した際に海上に残っ
たところで、アマミノクロウサギはここに生育していた薄暗い原始林地帯に
生息していたと述べている。ただ、本種は奄美大島全員に分布しているので
はなく、島の北部に位置する笠利町のように、標高が低く、原生林が存在し
ない地域には分布していない。アマミノクロウサギはこのような低標高地に
は元来分布していなかったのか、分布していたが、人為の影響で絶滅したの
かが定かでない。この点に関しては、歴史的に追跡する必要がある。
アマミノクロウサギは、奄美大島、徳之島に生育する原生林に好んで生息
しているが、昨今はパルプ材として原生林の伐採が進んでおり、分布域に変
動が生じている可能性が高い。また、1972年頃にはリュウキュウマツ、スギ
などの造林木に対する食害が両島の各所でクローズアップされていた。それ
は原生林の伐採跡地、即ち、アマミノクロウサギの生息圏内に植林が行われ
るためで、ここでも林木の生産と野生鳥獣との競合がみられる。早い時期に
アマミノクロウサギの存続のための保護区の設定などの方策が検討される時
期にきている。
第3回の調査では、奄美、徳之島両島で4地点から報告されていたが、今
回は7箇所からの報告がある。但し、2次メッシュでは徳之島の1箇所が減
少した。第4回の調査結果を見る限りでは、もっと広域に分布していると考
えられる。
アマミノクロウサギの消長に関しては、彼らの生息を許容している原生林
伐採との関連を調査する必要がある。近年、激しい森林伐採が行われている
ので、それがどのような形で影響を及ぼしているかを明らかにするための詳
細な調査が必要である。アマミノクロウサギは草食動物であるから、森林伐
採跡地に生育する栄養豊富な植生を餌に、一時的には個体数を増加させるが、
繁殖や休息といった生活の基盤として欠くことのできない森林がなくなるこ
とによって、一気に減少を始めることは十分に考えられる。
(2) エゾユキウサギ : B やや情報不足
(今回調査で使用した鳥獣保護員用調査票で、分布情報源である鳥獣関係統
計がユキウサギやカイウサギを区別せず、ウサギについてはノウサギとして
取り扱っているため、北海道に自然状態では分布しないノウサギのみが調査
票に記載されており、このためエゾユキウサギ(日本産唯一のユキウサギの
亜種)をノウサギとして報告したと考えられるデータが多数北海道に見られ
る。よって、今回ユキウサギとして報告された91メッシュのみでは、やや情
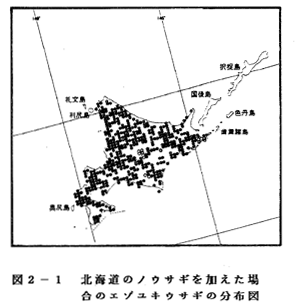
報不足気味であるが、北海道のノウサギの2,391メッシュをエゾユキウサギと
して加えると2,482メッシュとなる(図2-1)。これにより、分布状況は「
A 分布パターンを表わしている」に変る)
エゾユキウサギは北海道に
のみ分布することが知られて
いるが、その周辺に散在して
いる利尻、礼文、焼尻、奥尻、
渡島大島、同小島などの離島
からの生息報告はない。第3
回の調査では、日高、大雪両
山系のほか、根釧地方から40
メッシュの報告があったに過
ぎないが、今回は91メッシュ
の報告があった。前回に比べ
て、札幌から旭川にかけての
鉄道網、道路網に沿って顕著
に増加している。これは調査
者の分布や往来の頻度と深く
かかわりがあると考えられる。
しかし、全道的分布種と考え
られるところから、まだ情報
が不足していると思われる。 |
 |
古い分布情報がないので、分布域の拡大、縮小について論じる訳にはいか
ないが、環境庁発行の鳥獣統計関係(1923〜1986)によると、1959年には81
,860個体の捕獲がみられたが、これをピークにその後は徐々に下降し、1984
年には4,496個体にまで減少した。この原因は明らかではないが、毛皮とし
ての価値の下落やエゾユキウサギの有力な捕食者であるキタキツネの増加も
一因ではないかと考えられる。この捕食者と被食者の数の関係を鳥獣関係統
計でみると、図2-2のようになり、両者の間に相関関係がみられた(n=39、
p<<0.01)。
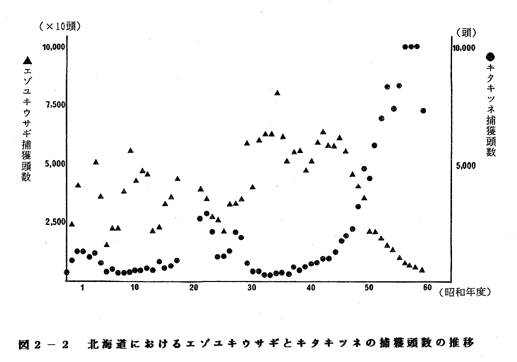
今回の調査ではエゾユキウサギもキタキツネも細部地域にわたる分布情報
がないので、地域別の相互関係は明らかではない。しかし、今後情報が増え
ればこの関係はより明確になるはずである。
キタキツネは北海道に多い風土病であるエキノコックスの媒介者として駆
除の対象となっており、今後の個体数の変動と、被食者であるエゾユキウサ
ギの個体数の変動を追跡していく必要がある。
(3) ノウサギ : A 分布パターンを表わしている。
ノウサギは北海道と沖縄県を除く地域に分布しているが、隠岐にはオキノ
ウサギが、佐渡にはサドノウサギが、本州、四国、九州にはトウホクウサギ
とキュウシュウノウサギが各々分布している。前回の調査では、鳥獣関係統
計その他の情報により生息が確認されているにもかかわらず、情報不足から
四国、佐渡、隠岐の各島からの分布情報は全くなかった。また、前回の調査
では104メッシュであったものが、今回は沖縄を除く全都道府県から報告が
あり40,602メッシュに達した。しかし、これは分布域が拡大したのではなく
て、調査精度が向上したに過ぎない。環境庁発行の鳥獣関係統計(捕獲個体
数)と、各県のメッシュ数の分布とを対比した結果、よく一致したことから、
分布調査は実態を反映しているものと考えられる。
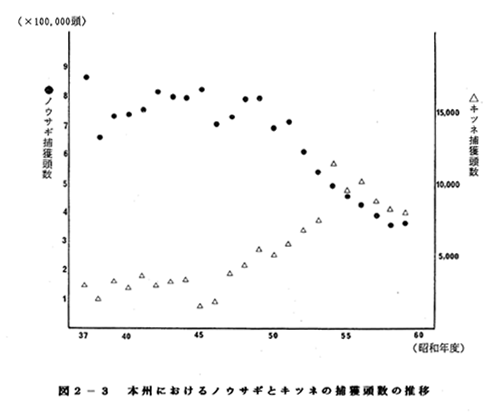
ノウサギとキツネの捕獲頭数の推移を図2-3に示した。この場合でも北
海道と同様に両者の間に相関がみられた(n=23、p<<0.01)。
サドノウサギについては、豊島ら(1970)の報告がある。
(4) カイウサギ : B やや情報不足。
本種は元来わが国には分布しないウサギであるが、毛皮動物、実験動物、
ペットその他の目的で外国から移入されたものである。前回は北海道と宮城
県から計5メッシュが報告されている。2次メッシュでは鹿児島県の島嶼で
も記録されている。実際には北海道の渡島大島、渡島小島(1984年7月20日
放獣)、岡山県の瀬戸内海の茂床島に生息していることが知られている(豊
島ら、1968)。また、奄美諸島のさる小島にも放獣されているともいわれて
いるが明らかではない(柴田義春、私信)など、まだ完全な記録とはいえな
い。
本種が離島に放獣された理由は明らかではないが、おそらく毛皮や肉を必
要とした時代にこれを目的として放飼されたものと思われる。渡島大島で筆
者らが1966年に調査した結果では、野生化した多数のカイウサギが元来貧弱
な島の植生を食い荒している上に、地中に穴を掘って生活するカイウサギと、
同じく地中に穴を掘って営巣するオオミズナギドリとが生活の場をめぐって
競合し、オオミズナギドリが全く営巣できないという事実があった。また、
地中に巣食うドブネズミも多く、オオミズナギドリの卵を食い荒し、営巣を
阻害していた。天敵不在の新天地に移入された鳥獣が数を増やし、自然植生
を破壊したり、在来の野生鳥獣を絶滅させた例は多い(Temple,1978)。
4-2. ネズミ目
(1) リス科
- (1) タイワンリス : A 分布パターンを表わしている。
本種は1933年に台湾から伊豆大島の動物園に移入された動物であるが、こ
れが逃げ出して数を増やしたものである。
このところ急速に分布域を広げ、第3回調査時には31メッシュであったも
のが、今回は623メッシュに増加しており、分布域が急速に拡大している。
これが実際に増えているとしたら、ニホンリスとの競合という見地から由々
しきことである。
東京都町田市の福祉施設と茨城県にあるさる霊園では、伊豆大島産のタイ
ワンリスを人間が入れる大きな檻の中で飼育、展示している。また、埼玉県
の大宮市では、エゾリスを放獣する計画がある。過去の事例から帰化種の多
くは動物園、公園などで飼育していたものが逃げ出して増加したケースが多
いことを考えると、動物の移動に対して何らかの法的規制が必要と考える。
静岡県伊豆半島でもタイワンリスが増加し、電話線を切断する被害が続出し、
対策協議会を発足させて対策を検討中である。秋田県、岩手県の県境付近の
十和田八幡平国立公園付近、宮城、秋田、岩手の県境付近の栗駒国定公園の
観光地にも出没した。内陸の1,000m前後の高標高地に出没していることか
ら、ニホンリスとの競合が懸念される。これまでの調査で伊豆大島と瀬戸内
海の友ヶ島、瀬戸内海の沖ノ島、神奈川県の江ノ島、鎌倉、東京の新井薬師、
石神井公園、姫路城公園、神戸のロックガーデンにおける生息が知られてい
る(宇田川、1954、朝日、1967、尾崎、1986)。
本種が動物公園から逃げだした事例を除いて、各地に分布しているものが
どのような経緯で出現したかは不明であるが、鎌倉ではペットとして給餌し
ているタイワンリスが樹皮を噛り枯損させるという弊害が出ている。また友
ヶ島では樹木に相当な被害が出ているという(朝日、1967)。
一方、タイワンリスと類似した生態的地位を持つニホンリスとの競合から、
在来種を駆逐する危険性も高く、移動の制限、現存個体の除去などの対策が
望まれる。鳥獣関係統計によると、東京都では毎年1,000〜2,000頭前後のリ
ス類が捕獲されていることになっているが、これらの中には伊豆大島におけ
るタイワンリスが大きな比率を占めている。東京都が狩猟者に対して行った
アンケート調査によると、昭和60年度の全捕獲数1,196個体のうち99.5%に当
たる1,190個体が伊豆大島で捕殺されたタイワンリスである。今後、東京都
以外の府県でもタイワンリスの分布拡大の動向を知る手段として、狩猟また
は有害獣駆除、或は学術研究用として捕獲されたものについては、報告を義
務づけることが望ましい。
(2) エゾリス : A 分布パターンを示している。
本種は北海道にのみ分布する種であるが、それに付随する島嶼からの報告
はない。第3回調査時には14メッシュであったものが、今回は1,891メッシ
ュに増加している。分布図の上から見る限り飛躍的増加に見えるが、調査精
度が向上したに過ぎない。今回のメッシュ数はほぼエゾリスの分布の実態を
表わしていると考えられる。
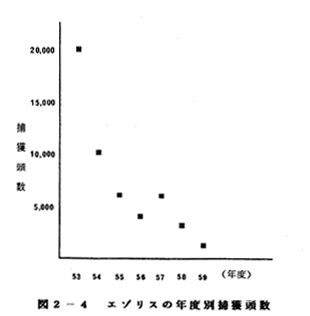
鳥獣関係統計では、
狩猟による捕獲数は
1980年には21,422頭
であったものが、
1986年には1,299頭
にまで激減しており、
何らかの保護対策が
望まれる(図2-4)。
本州の調査者の中
にエゾリスと記した
人がいるが、埼玉県
大宮市でエゾリスを
放獣する計画がある
ことから、本来の分
布域である北海道以
外の地域に存在する
可能性はある。 |
 |
(3) ニホンリス : A 分布パターンを示している。
本種は北海道を除く本州、四国、淡路島、九州に分布していることが知ら
れている(今泉、1960)。第3回調査時には72メッシュであったが、今回は
9,527メッシュに増えた。この飛躍的増加は、分布域の拡大ではなく調査精
度の向上と見るべきである。
前回の分布調査では、宮崎、熊本、鳥取、島根、岡山など34県でゼロであ
ったが、今回は、もともと分布しない沖縄を除いて、鳥取、長崎、鹿児島の
3県から記録がない。この3県は前回もゼロであった。このうち宮崎、熊本
両県は狩猟による捕獲がゼロなのに、今回の調査で出現しているし、鹿児島
は1986年に18個体、1989年には2個体が捕獲されている。これがニホンリス
であるのか、タイワンリス、シマリスであるのかを確認する必要がある。仮
に、ニホンリスであるとしたら、保護のために狩猟獣から除外して回復を図
る必要がある。いずれにしても西日本一体では、前回の調査時と同様に希薄
な分布パターンを示している。従って、このような地域においてこそ経年的
な分布動向を知る必要があると同時に、その結果に基づいた対策を立てる必
要のある地域ということになり、この意味から詳細な情報が望まれる。タイ
ワンリスの項でも触れたが、ニホンリスとの競合の結果、ニホンリスの分布
域の縮小が予想されることから、両種が接する地域での情報収集は5年周期
よりも短いことが望ましい。また、このような地域では年々の分布域の変動
を記録することは意味のあることである。
(4) シマリス : A 分布パターンを示している。
わが国には北海道にのみ分布する在来亜種のエゾシマリスと朝鮮半島など
からペットとして移入されたチョウセンシマリスが篭抜けして生息している。
しかし、移入されたチョウセンシマリスについては情報不足からその実態は
明らかではない。
前回は北海道の26メッシュ以外はどの県もゼロであったが、今回は北海道
で923メッシュ、16都県で220メッシュが報告されている。シマリスは千葉
県の野田市、山梨県の鳴沢村の富士山麓で観察されているが、千葉県からは
これに関する報告がない。いずれにしてもこれほど多くの地域に外来種が出
現していることは、動物地理学的に由々しいことである。特に、北海道にお
けるシマリスの分布が、北海道のエゾシマリスなのか、朝鮮半島などから移
入された外来種であるのかを確認する必要がある。北海道に外来種が侵入す
ると、近縁種であるエゾシマリスとの交雑が懸念される。
今回の調査でも、シマリスとして情報の提供を求めたという経緯から、上
記2種の明確な識別はなされていないが、北海道以外のシマリスは外来種で
あると見てよかろう。しかし、北海道のシマリスは、在来種なのが外来種な
のかを明確にする意味において、詳細な調査をする必要がある。
チョウセンシマリスの輸入の実態は明らかでないが、これまでの情報では
札幌の北大植物園内に過去9年間にわたり外来種のシマリスが放獣されてい
たという。本州方面では、山梨県南都留郡鳴沢村の富士山麓にあるスバルラ
ンド森林公園から逃げ出したと思われるシマリスが生息しているほか、新潟
県の弥彦山、角田山にも生息していたという(風間辰夫、私信)が、その後
のことは不明である。
(5) ホンドモモンガ :B やや情報不足。
本種は本州、四国、九州などに広く分布しているが、夜行性であるところ
から人目に触れる機会が少ないので情報数が少ない。前回は3県から13メッ
シュが報告されていたが、今回は9県から24メッシュが報告された。いずれ
も愛知県以北からである。
本種は狩猟獣でないところから、当調査以外の資料による情報収集が困難
であるが、まだまだ情報不足による空白県が多いと思われる。
(6) エゾモモンガ : B やや情報不足。
本種は北海道にのみに生息する種である。前回は7メッシュであったが、
今回は13メッシュに増加した。しかし、既往の知見からいって正確な情報が
得られているとは考えられない。今回の調査でも石狩、十勝、網走、根室地
方からわずかに報告があるが、もっと広域に分布していると考えられるので、
より一層の情報収集に努めるべきである。ホンドモモンガと同様、非狩猟獣
であることから、資料が乏しく考察が難しい。
(7) ムササビ : A 分布パターンを示している。
本種は北海道を除く本州、四国、九州に分布しているが、生息する地方に
よってニッコウムササビ、ワカヤマムササビ、キュウシュウムササビの3亜
種に分類されている。
前回は12都県から53メッシュが報告されているが、今回は千葉県、佐賀県、
長崎県の3県を除く43都府県から4,400メッシュが報告されている。
前回の調査では、中国、四国方面からは皆無であったが、今回は報告があ
った。従って、今回報告がなかった3県についての詳細な調査が望まれる。
本種は夜行性であるが、独特の鳴き声と、夜間滑空することから比較的人目
につき易いこともあり、夜行性にしては目撃情報の多い動物といえる。
(2) ヤマネ科
(1) ヤマネ : B やや情報不足。
本種は1科、1属、1種の日本固有種で、北海道を除く本州、四国、九州
地方に生息している。前回は青森、長野の2県から20メッシュが報告された
が、今回は11県から50メッシュが報告された。京都府を始め他県にもいる
はずであるが、今回の調査では極(限られた情報しか得られていない。夜行
性である上に冬眠するという不利な条件により情報が少ないものと考えられ
る。
-
- (3) ネズミ科
- (1) ミカドネズミ : A 分布パターンを示している。
本種は北海道にのみ分布する種で、北海道内では比較的広く分布すると考
えられている。前回には72メッシュであったものが、今回は情報量が多くな
り114メッシュに増加した。それでも道北と道南部からの情報が少ない。こ
れは研究者の分布の偏りや採集の有無を反映しているように思われる。しか
し、詳細は不明である。
(2) エゾヤチネズミ : A 分布パターンを示している。
本種は北海道にのみ生息する種で、本州におけるハタネズミと同様に若齢
造林木を食害するところから、その被害防止のための研究が良く行われてき
た。従って、ネズミの仲間でも分布や生態が非常に良く解明されている種と
いえる。
前回は123メッシュであったが、今回は201メッシュに増加し、情報量が多く
なった。
(3) シコタンヤチネズミ : A 分布パターンを示している。
北海道にのみ分布する種である。前回は利尻島から2メッシュのみが報告
されていたが、今回も同じく2メッシュで変化がない。種の特異性や分布の
地理的特性から、これ以上の大幅な情報収集は望み薄である。本種の実態は
不明な点が多い。
(4) リシリムクゲネズミ : A 分布パターンを示している。
北海道北部の利尻、礼文両島に分布していることが知られている。( Ima-
izumi、1971)が、両島内における分布や個体数に関する情報は乏しい。
前回は5メッシュであったが、今回は12メッシュになった。北海道本島内
にも分布が見られるようになった。
(5) ミヤマムクゲネズミ : A 分布パターンを示している。
本種は北海道の日高山系からの分布情報があるが、これまでも詳細な分布
調査が行われていないので.分布の精度などについては考察できない。
前回の調査と今回の調査でまったく同じ18メッシュが報告されている。
(6) ニイガタヤチネズミ : A 分布パターンを示している。
本種は本州の中央部から分布情報が寄せられているが、これまでの知見で
も、本州中部及び関東地方の高山及び亜高山となっている。前回は30メッシ
ュであったが、今回は44メッシュが記録され、かなり増加した。今回の分布
情報は妥当なものと考えられる。
(7) トウホクヤチネズミ : A 分布パターンを示している。
本種は青森、岩手、福島の各県に分布していることが知られている。前回
は13メッシュであったが、今回は60メッシュに増えた。紀伊山地から2例の
情報が寄せられているが、これまでの採集例も少なく、詳細は不明である。
(8) ワカヤマヤチネズミ : A 分布パターンを示している。
前回は和歌山県と奈良県から各1メッシュが記録されたが、今回は同じく
両県から6メッシュが記録された。
(9) カゲネズミ : A 分布パターンを示している。
本種は関東及び中部地方に分布していることが知られている。
前回は5県から3メッシュが記録されたが、今回は7県から48メッシュが記
録された。しかし、既存の知見を下回る情報しか得られていない。
(10) スミスネズミ : A 分布パターンを示している。
本種は富山県以西の本州、四国、九州及び隠岐島に分布していることが知
られている。前回は8県から18メッシュであったが、今回は22府県から85メ
ッシュが記録された。最も北では福島県から2メッシュが記録されたが、分
布域かどうか確認する必要がある。今回の調査では隠岐島からの報告がない
ほか、本州西部で欠如している県もあり、より一層の努力が必要である。
(11) ハタネズミ : A 分布パターンを示している。
本種は本州及び九州に広く分布していることが知られているが、今回の調
査では分布情報が極端に片寄っている。ハタネズミは北海道におけるエゾヤ
チネズミと同様に造林木を食害するため、分布、生態に関する調査は他のノ
ネズミに比べて良く行われているが、その割には情報提供が少ない。佐渡に
も分布していることが知られている(宮尾ら、1968)。
前回は67メッシュであったが、今回は143メッシュに増加した。しかし、
西日本には分布が希薄である。
(12) マスクラット : C 情報不足。
本陣は北米産であるが、毛皮として日本に輸入された後、逃げだしたり、
毛皮価格の低迷で放逐されたものが野生化し、1947年頃から東京都江戸川付
近でみられるようになったほか、埼玉県でも捕獲された例がある。
今回の調査では1件の情報も見られなかったことから、絶滅したのではな
かろうか。
(13) アカネズミ : A 分布パターンを示している。
本陣は本州、四国、九州、壱岐、五島に分布していることが知られている。
北海道を含む佐渡、伊豆大島などの島嶼のものはそれぞれ別亜種とされてい
る。
前回は269メッシュであったが、今回は635に増加した。本種は世界的規
模で分布しており、わが国でも全国的に分布していることが知られているが、
その割には分布情報が少ない。
(14) カラフトアカネズミ : A 分布パターンを示している。
北海道に分布することが知られており、今回の分布情報は比較的妥当なも
のと考えられる。前回は31メッシュであったものが、今回は49メッシュに増
加した。
(15) ヒメネズミ : A 分布パターンを示している。
本種は全国に広く分布しているが、島に分布しているものは各々別亜種と
されている。前回の調査では206メッシュから今回は506メッシュに増加し
た。これでも情報数が少なく、完全な分布図とはいえない。
(16) カヤネズミ : C 情報不足。
本州の関東以西に広く分布していることが知られているが、今回の調査で
は西日本で情報量が少ない。前回は31メッシュであったが、今回は74メッシ
ュに増加した。本種は淡路島にも生息していることが報告されている(宮尾
ら、1983)が、淡路島から依然として報告がないなど、まだまだ十分な分布
域を表しているとは限らない。
(17) ハツカネズミ : B やや情報不足。
本種は日本全国に分布していることが知られているが、島嶼に生息するも
のはそれぞれ亜種に分けられている。
前回は44メッシュであったものが、今回は101メッシュに増加したが、25
府県から報告がないなど、十分に分布域を表わしていない。今回の調査でも
総体的に分布情報に乏しいが、徳之島、沖縄本島、小笠原などの島嶼からの
情報の多いことが特徴である。淡路島からの分布情報もある(宮尾ら、1983)。
(18) オキナワハツカネズミ : A 分布パターンを示している。
奄美大島と徳之島の両島から2メッシュが記録されている。
(19) アマミトゲネズミ : A 分布パターンを示している。
奄美大島と徳之島の両島から5メッシュが記録されている。
(20) クマネズミ : C 情報不足。
本種は全国的分布種である。前回は北海道と本州から18メッシュが記録さ
れたが、今回は34メッシュでやや増加した。本種はほぼ日本全国に分布して
いると思われるのに空白箇所が35府県に及んでいる。これまでに西表島、沖
縄本島、小笠原諸島などから報告があったほか、八丈島(今泉、1967)と鳥
島(Yabe,1982)からも記録がある。
本種は人類の移動に伴って世界各国に分散した種であるが、ドブネズミと
共に島嶼に上陸して在来種を捕食し、大きな打撃を与えることが知られてい
る。セイシェルメジロは侵入してきたネズミ(Ruttus sp.)によって殆ど絶
滅させられたと考えられている(Temple.1978)。
ドブネズミ : B やや情報不足。
本種は全国的に分布していることが知られている。
前回は60メッシュであったものが、今回は91メッシュに増加したが、空白
地帯が27府県もあるなど必ずしも実態を反映していない。この現象はクマネ
ズミなどの普通種でも共通した傾向を示している。このような種は、今後、
一般国民に参加を呼びかけても、クマネズミとの識別が困難なので、実態に
即した情報を得るには研究者の協力に頼る以外に方法がなかろう。
ケナガネズミ : A 分布パターンを示している。
前回は報告がなかったが、今回は3メッシュの記録があった。
(5) ヌートリア科
- (1) ヌートリア : A 分布パターンを示している。
ヌートリアは明治末期にわが国に移入されたといわれている(三浦、1976)
が、詳細は不明である。その後、毛皮獣として本格的に移入されたのが1930
年代といわれ、今では飼育されていたものが逃げ出したり、毛皮価格の低迷
で放逐されたりして野生化している。
前回は全く記録がなかったが、今回は1,460メッシュと急増した。これは分
布域を拡大したのではなくて、調査の精度が向上したものと考えられる。
本種は農業被害、例えば、そ菜類の食害、水田の畔に穴を掘るなどの被害
が各地で報告されており、有害獣として1963年以降、狩猟獣に指定されてい
る。鳥獣関係統計によれば、岡山、広島、兵庫、京都、神奈川などで捕獲記
録がある。今後、分布の動態を把握していく必要がある。
4-3. むすび
ウサギ目、齧目の分布調査結果を通じての特徴の一つは、情報が偏在してい
ることである。例えば、ノウサギ、ニホンリス、ムササビなどに見られるように
集まった情報を見ると、長野、宮城、東京、千葉、愛知の各県に集中しており、
日常茶飯事に遭遇しているであろう府県からの情報が少なく、情報源の偏在傾向
が著しい。
前回調査時には9府県からは、他の哺乳類も含めて1件の情報提供もなかった
が、今回はさすがにこのような無情報府県はまったくなかった。それだけこの調
査がこれまで無関心であった府県を啓蒙したことになる。調査結果が片寄ってい
るのは、調査を担当した研究者の分布や研究者が持つ調査地の分布とよく一致し
ているように思われる。今後、全国的規模で精度を上げて行くためには、まず情
報源の拡大を図らなければならない。これに反して、対象種が特殊なものや、分
布が特定の島嶼やどく限られた地域の場合、例えば、ナキウサギ、タイワンリス、
ミヤマムクゲネズミなどは、比較的情報数も多くかつ確度の高い情報が得られて
いる。これは研究者の関心が高いため調査がよく行われ、報告書が出ているため
と、古くから既知の事実となっており、分布域の把握が容易なためと考えられる。
これに対し、極く普通に数多く分布しているノウサギ、ユキウサギ、ニホンリス
などは意外と情報量が少ない。これら普通種の情報数を増やすためには情報源の
拡大を図る以外にない。
今回の情報提供者となった研究者が今後調査地を拡大して行かない限り、この
形式の情報収集を図っても、飛躍的に情報量が増加することは期待できない。従
って、今後は信頼のおける小中高の教員を始め、博物館、郷土資料館等への協力
要請も検討すべきである。今後、当調査が継続されるに従って、特定の種につい
ては順調に情報数が増えるのに対し、ハツカネズミのような研究者の関心の薄い
普遍種の情報は集まりが悪いものと考えられる。例えば最も身近に住んでいるド
ブネズミ、クマネズミであっても専門の研究者以外の人が目視によってこれを識
別し、情報を提供するという可能性は極めて薄いことから、この方面の情報数を
増やし、精度を高めるにはどうしても研究者の協力が必要である。従って、上記
3種を含めたネズミの仲間の分布図作成の成否は、いかに研究者の協力を取り付
けるかにかかっていよう。
前回の調査で、齧歯目ネズミ料のアマミトゲネズミ、ヌートリア科のヌートリ
アなどは現存するにもかかわらず、情報提供が皆無であったが、今回は情報提供
があった。これは今回の調査の精度の向上を示しているといえよう。
このたびの一連の全種を対象とした分布図作成の試みは、わが国では最初のこ
とでもあり性急に多くを望むことは無理であるが、この試みはわが国の野生鳥獣
の合理的な保護並びに管理を推進する上で極めて重要な資料を提供するものであ
り、今後とも、より積極的に取り組む価値がある。
そこで本調査をより効率的に進めるための手段として、過去に出版された種々
の文献や資料を引用することを提言したい。今回担当したウサギ目と齧歯目の考
察を行うに当たり、身近な文献をひもといただけでも、今回の調査を補完するデ
ータに数多く遭遇することが出来た。文献は学術誌に限らず、各都道府県の刊行
物、博物館、資料館等の出版物、自然保護協会等の財団による出版物、さらには
開発事業非体の行うアセスメントのための調査報告書等々、有用な資料は数多い。
次回の調査実行までの間に全国的規模での資料の収集、解析を実施すると共に、
それらの調査資料のデータベース化と集中管理するシステム(データバンク)の
確立を提言したい。
さて、帰化種による在来種への影響は、カイウサギの項で一部述べたが、わが
国で帰化種のいる場所として、小笠原諸島父島、兄島、ムコ島、八丈島、八丈小
島等のヤギ、宮城県金華山のニホンジカ、本州西部、九州地方のチョウセンイタ
チなどが知られている。わが国の野生生物保護上、動植物のみだりな移動を制限
する何らかの法的措置が望まれると共に、帰化動物の分布とその影響の実態把握
を早急に行う必要がある。
奄美群島、沖縄、小笠原、伊豆諸島などの島嶼群は、古くに大陸から孤立した
こともあって、生物が極めて特異的な進化を遂げ固有種も多い。これらの島々の
生物相は、気候的、地史的あるいは生物の相互佐用によって、長い歴史的過程の
中である種の安定を保ってきた。しかし、近年、森林伐採、道路網、観光施設等
の整備、人口増、外来種の進入などにより、ただでさえ脆弱な生物相は有史以来
の試練を迎えている。例えば、奄美大島における森林伐採、奄美大島、西表島に
おけるマングースの帰化、三宅、八丈島におけるイタチの放獣等々枚挙に暇がな
い。このような生息基盤の破壊、強力な補食者の侵入は対抗の術を持たない島嶼
の生物に大きな打撃を与えている。しかし、これらの島嶼における生物調査は1
〜2の島嶼群を除いて全く行われていないか、10年、20年の昔に行われて以来、
記録がないなど実情を知る上で極めて憂慮すべき状況にある。今回の一連の調査
結果でも、島嶼における情報量が本州、四国、九州、北海道などに比べて相対的
に少ないことが浮き彫りにされた。かかる実状を踏まえて、今回の自然環境保全
基礎調査とは別個に、これら島嶼群の生物相を対象とした緊急かつ系統的な調査
が望まれる。
( 阿部 學 )
引用文献
朝日稔ら、1967.友ヶ島のタイワンリス.日本哺乳動物学会誌.3(6):152-157.
芳賀良一、1957.ナキウサギの生態と林木被害.北方林業.9(11):334-336.
今泉吉典、1960.原色哺乳類図鑑.保育社.196pp.
今泉吉典、1967.住家性クマネズミの分類.特に八丈島の亜種について.日本哺
乳動物学会誌.9(2):128-143.
IMAIZUMl,Yoshinori,1971.A new vole of the Clethrionomys rufocanus
group from Rishiri Island,Japan. J. Mamm. Soc. Japan. 5(3):99-
103.
環境庁、1923-1986.鳥獣関係統計.野生生物課.
桐野正人、1977.アマミノクロウサギ.汐文社.214pp.
三浦慎吾、1976.分布からみたヌートリアの帰化・定着.岡山県の場合.日本哺
乳動物学会.6(5/6):231-237.
宮尾蝦雄ら、1968.佐渡における小哺乳額の採集結果.日本哺乳動物学会誌 .
4(1):29-32.
宮尾嶽雄ら、1983.淡路島南部の噸乳類.日本哺乳動物学会誌.9(2)=128-143.
日大農獣医学部、1969.小笠原諸島の調査.日大農獣医学会誌.1969(3):12-51.
尾崎研一、1986.タイワンリスの植物と菜食行動.日本哺乳動物学会誌.
11(3/4):165-172.
TEMPLE.Stanley A..1978. Endangered Birds. Univ.of Wisconsin Press.
豊島重造ら、1968.野生化した家兎の生息数の推定と生態に関する2、3の試み
と考察.新大農園報3:1-5.
豊島重造ら、1970.サドノウサギの生態に関する収集資料の検討.新大農園報.
4:69-76.
内田映、1960.北海道におけるナキウサギの分布.北方林業.12(7):235-239.
宇田川竜男、1954.伊豆大島におけるタイワンリスの生態と駆除.林試研報.
67:93-102.
YABE.Tatsuo. 1982. Habitats and habits of Roof rat. Rattus rattus on
Torishima.tha lzu lsland. J. Mamm. Soc. Japan. 9(1):20-24.