�@
�P�D�����̖ړI�ƕ��@
�������z�����̈�Ƃ��čs��ꂽ�����ޕ��z�����́A�\�Q�|�V�|�P�Ɍf����10��́u�w�W�����ށv���\�Q�|�V�|�Q�Ɍf������ɂ���ēs���{�����ɑI�肵���u���荩���ށv�ɂ��āA�����n�̈ʒu�A�������Ɛ����̊T���A�ی�̌�����������̂ł���B
�u�w�W�����ޕ��z�����v�́A�䂪���ɐ�������10����ȏ�̍����ނ̂����A���z�悪�L���A��r�I����݂�����A���S�̂Ƃ��ĎR�n���畽�n�܂ł̗ǍD�Ȏ��R���̎w�W�ƂȂ鍩�����w�肵�A���ׂĂ̓s���{���ɂ����ď�L�̍��ڂ��������ꂽ�B
�{�����̑Ώێ�̐������́A�R�n���畽�n�Ɏ���܂ł̉�������ς��i��ł��Ȃ������i�k���A����A���n�A�r���j�⎩�R�т�тȂǂŁi�\�Q�|�V�|�R�j�A�����͐l�Ԃ̐�����̒��ɂ݂���ǍD�Ȏ��R���A�����闢�R�E�c���I���R�̑�\�I�v�f�Ƃ݂Ȃ�������̂ł���B
���������āA�{�����̎��{�ɂ���āA�Ώێ�̕��z�̔c�����\�ƂȂ�ق��A�ǍD�Ȏ��R���̑S���I�ȏ���������x�c�����邱�Ƃ��ł���Ɨ\�����ꂽ�B
�u���荩���ޕ��z�����v�́A�������@���͎w�W�����ޒ����Ɠ���ł��邪�A�����Ώێ�͗\�ߒ�߂��Ă��炸�A�e�s���{����50�`100���I�肵�A������Ώۂɍs��ꂽ�B
�I�����I�m�ɓK�p�����A��ł̊댯���̒��x��w�p��̏d�v�����̊ϓ_����A��̈ʒu�Â������̂����疾�炩�ƂȂ�A���ۑS�̏��{��̒��ɁA�����ނ̕ی����������\���������Ă��悤�B
�{�����̒��������́A�����n�̈ʒu�A�������A�����̊T�v�A�ی�̌��ł���B
�����͓��{�����w��̋��͂āA��������������𒆐S�Ƃ��A�K�v�̂���ꍇ�ɂ͌��n�����A�����݂Ȃǂɂ��A�\�Ȕ͈͂Œm�������W�����B
�u�����n�̈ʒu�v�͍��y�n���@���s��20�����̂P�n���}�ɂ�������ŕ\�����A�����n���������ꍇ�͏����ۂŕ\�������B
�u�������̌���v�́A�����n���Ƃ̐������̌���ɂ��āA���\�̊�̂�����̋敪�ɊY�����邩�������B
|
�ǍD |
���������ǍD�ɕۂ���Ă���B |
|
�s�� |
���������������B |
|
�j�� |
���������j��āA���Y�����������ł��Ȃ��Ȃ��Ă���B |
�u�������v�́A���Y�����n�ɂ����铖�Y��̐����������܂��ɔ��f���āA���\�Ɏ����敪�̂�����ɊY�����邩���肵�A�Ή�����L���ŕ\�����B
|
�Ȃ��A�����╷�����ݓ��Ő�������Ƃ�����������ꍇ�ł��A�����_�ɂ����āA�����ɂ͐������Ȃ��ƒ����҂����f�����ꍇ�A�܂��Ē�����K�v�Ƃ���Ɣ��f�����ꍇ�ɂ͂��̎|���L���ꂽ�B
�Â��L�^�ŁA�Y�n�����ĂłȂ����́A�Ⴆ�Δ��R�Ƃ����n���⑭�̂��p�����Ă���ꍇ�ɂ́A�n���}��ɐ����n���������Ƃ͍s���Ă��Ȃ��B
�������ʂ͕��z�}�ƒ����[�ɕ����ċL�^����A���z�}�́A���y�n���@���s��20�����̂P�n���}�ɁA�w�W�����Ɠ��荩���ɕ����ĕ\�����ꂽ�B�������A�����n�����\�����Ɨ��l�̋��ꂪ�����ɂ��ẮA�����҂̔��f�ɂ��A�L�ڂ���Ă��Ȃ��ꍇ������B
�����[�͓s���{���ʂɂ܂Ƃ߂��A�n�悲�Ƃ̊T����t���ĕ��Ƃ��Ĉ�����ꂽ�B
�����ނ̕��z��S���I����Ŕc�����邱�Ƃ�ړI�Ƃ��āA���f�[�^�̋L�ڏ��̎��C�e�[�v����ƂƁA�w�W�����ɂ��Ă̕��z���b�V���}�̍쐬�A�d�Z�@���g�p�����������̏W�v��͂��s�����B
����̒����͏�������A���̓��e�����z���}�ƒ����[�̂Q�ɕ�����Ă������߁A�W�v�E��͂ɓ������Ă͗��҂̏ƍ����K�v�ł���A��v�Z�ł͑���ȘJ�͂�K�v�Ƃ��邱�ƁA����Ɏ����̕ۑ���̗��_���l�����āA�����e�̎��C�e�|�v���͉����s�Ȃ����B
���͂ɓ������ẮA�����[���e�A���z���}�L�ړ��e���\�Ȍ��萳�m�ɓ��͂���悤�w�߂��B���ɕ��z���}���e�̓��͂ɓ������Ă͂ł��邾�������ȒP�ʂœ��͂��邱�ƁA�W�v�E��͂ɂ������đ��̊֘A���Ƃ̂��荇�����\�ł��邱�Ɠ����l�����Ċ���b�V���i�O�����b�V���j��P�ʂƂ��ē��͂����B
���͕͂��z�}���ƒ����[�������ꂼ��ʁX�ɍs���A�d�Z�@�����ɂ���ĂQ�̏��������t�@�C�����쐬�����B
�{�����̏ꍇ�A�ΏۂƂ����킪���v1,764��̑����ɂ̂ڂ�A�����̌��ŋ��ʂ��Ē������ꂽ�킪���ɏ��Ȃ��������Ƃ���A��L�̕��@�ɂ���č쐬���ꂽ���C�e�[�v��p���Ă̏W�v�́A�S�����ʂɒ������ꂽ�w�W����10��𒆐S�ɍs���A���z�}�̍쐬���w�W�����݂̂Ɏ~�߂��i�t�\�P�y���}�Q�|�V�|�P�`10�j�B
���荩���ނɂ��ẮA���ψ���ݒu����A�a���A�w���̌�����I���̓K�����̌������s��ꂽ�B
�@
�@
�@�����ޕ��z�����ɂ����Ďw�W�����ނƂ��đI�肳��A�S�����z���������{���ꂽ10��̍����ɂ��āA�����j�y�юw�W���A���z��l�דI�e���̎�ނƒ��x�Ȃǂ𒆐S�ɉ�������B
�@�{���́A��Q�R���ۑS��b�����A�������z�������i�����ށj�S���łɂ܂Ƃ߂�ꂽ��匤���҂ɂ���ʉ�����A�K�v���������ĕҏW�������̂ł���B
�@
�P�j���J�V�g���{�@Epiophlebia superstes
���J�V�g���{�ނ���x�ޒ��ł����قȍ����ŁA������ɐ����������J�V�g���{���ڂ̈⑶��ƍl�����A�����ł̓q�}�����n���ɂP��A���{�ɂP����Y���邾���ŁA�n���w�ォ����d�v�ȍ����ł���B�������䂪���ł͖k�C�������B�܂ŗ����������i�B��͗�O�j�S�哇�ɂ͊e���ɕ��z���A����̊����\����̂Ŏw�W�����Ƃ��ēK���ł���ƍl������B
�����͏t���o������B����{�ł͂R��������T���܂ŁA�k���{�ł͂T�E�U�������̊��߂ŁA�H����P�`�Q�T�Ԃ͗c���̐����n�ł������R�Ԃ̌k�J�ɉ��ĐېH�ɉ߂��A���n����ΗY�͓����k�J�ׂ̍��k���ɉ����āA�������㗬�ւƔ��Ă��A�������߂�B�����Y�͎���ɐÎ~���Č�����s�����A�����̌�Ɏ��͒P�ƂŎY���s���ɓ���B���͌k���̉��ɐ����Ă���t�L�E���T�r���̑��̏_��ȗt���E�s�����A���̑g�D�̒��ɎY�ݍ��ݗ���20���ł͖�30���ʂ���q����̌�ɛz�����A�c���͓��H���s�����A�����̐̊Ԃɐ������A��13��̒E������s���Đ�������B���̑S�������Ԃ͖��m�ɒm���Ă��Ȃ����A���炭�T�N�ȏ�V�`�W�N�܂łƍl�����Ă���B����̂��߂ɂ͐����E�ቷ�̗����K�v�Ƃ��A���n�c���͉H�����̑O�A���炭�P�`�Q�����O�Ƒz������邪�A����𗣂�ĕt�߂̊��I�̉��Ȃǂɓ���H����҂B���̊Ԃ͐����ċz�����C�ċz�ֈڂ�ς����̂Ǝv����B�H���͋ߖT�̒n���i��E�����Ȃǁj�ɓo���āA�̈ʂ𐅕��ʂɑ�110���`135���̊p�x�̓|���^�Ƃ��ĒE�炷��B
�{��͕��ފw�I�ɂ��`�Ԋw�I�ɂ����ق̍����ł���݂̂Ȃ炸�A���ʂ��e�ՂŁA�U�S���I�ɕ��z����̂Ŏw�W�����Ƃ��ċɂ߂ēK���Ă���B�X�ɐ����ɂ��c���ɂ�����������ꂽ�Ɠ��̊��A�������ɕ���ꂽ�}�Ζʂ̂��邹�܂��k�J�̐���ȗ����Ƃ��̏���̋�Ԃɐ������āA�R���{�킪�������Â��邽�߂ɂ͈ꐢ��V�|�W�N��v���邱�Ƃ���A���N���ɘj���Ĉ������ۑS����邱�Ƃ������Ƃ���B��������悭���R�̕ۑ����ꂽ�R�Ԍk���̐����̑�\�ƂȂ��Ă���̂ł���B�{��͂����ꂩ�Ɖ]���Β�R�n���̎�ŁA�C��100m�ʂ���1000m�ʂ܂ł̊ԂɌ�����B�����P���ߖT�̎Y�n1500m���O�͂ނ����O�I�ȕ��z�Ɖ]���悤�B
����̑S�������ɂ���āA�{��͑S��47�s���{���̂����R���i�{��E��t�E����j��������44�{������L�^���ꂽ�B����ɂ͒n���I�E�n���I�E�C��I�ɂ��������Ȃ����Ƃ������ł��邪�A�{�錧�ł͍�������������݂�ׂ��ł��낤�B��t���͊J���̐i��ł��邱�Ƃ�ʂƂ��Ă��A���J�V�g���{�̐����������ɏ\���Ȋ��̌�������R�n�����Ȃ����Ƃ�茩�āA����������̌����݂��Ȃ��悤�ɍl������B
�@
�Q�j���J�V�����}�@Tanypteryx pryeri
���̗ނ̃g���{�͓��{�ł̓��J�V�����}�P�킾�����Y���邪�A���ݐ��E�Ŗk�āE�`���[�E�I�[�X�g�����A�y�уj���[�W�[�����h�ɍ��v�T��10����Y���邾���ŁA�s�������ڂ̒��ōł����q�I�Ȍ`�ԂƓ��قȐ��Ԃ������A�⑶�����̈�ł���B����A�k�C���E��t�E�����y�юl���̂S���ƒ���E�����������36�{������L�^���ꂽ���A���������͍Ē�����K�v�Ƃ��邽�ߐ�Ŏ�ɓ�����Ă���B
�����͂T�E�U���Ɍ���邪�A���k�n���Ⓑ�쌧�ł͂�����ĂV��������W���͂��߂܂Ō�����B���������J�V�g���{�Ƌ��Ɍ����邱�Ƃ����邪�A��R�n�̖T�̓��H��Ȃǂ����������Ă��A�����Ђ炢�Ď�����A��A�t��A���H��A���ɂ͐l�̌��Ȃǂɂ��Ƃ܂�B������ߐH���Ă��邪�A��^�ŐÎ~���Ղ��̂Ŕ����e�Ղł���B
�Y���͊R���̟��o���̂��鎼�n�̐��ہE�y���ȂǂɒP�Ƃ̎������[��}�����čs���A�c���͂��̂悤�ȏꏊ�̓y���������͐��ۂ̉��̂��ڂ݂Ȃǂɐ���ł���B�y������ꍇ�ɂ͎��y���ɃJ�j�̍E�̂悤�ȃg���l��������A���̓����ɓ������O�Ɍ����č���A�ʉ߂��鑼�̏�������߂��ĐH�ׂ�B���J�V�g���{�̂悤�ɐ����}�����ɐ�������̂ł͂Ȃ����A���o���͉�������Ă͂Ȃ�Ȃ��B�c���̗���y�їc�����Ԃ̒����͐��m�ɂ͒��ׂ��Ă��Ȃ����A���炭���n����܂łɂS�`�T�N��v���āA12�`13��̒E����s�����̂ł��낤�B
���n�c���͒�n�ł͂S�����{����T����{�Ɏ��n�̑��ȂǂɂƂ܂��ĉH������B���Y�n�̎��n�ł͑����̒E��k���Ⴂ�A���̂ɂ��Ă���̂����邱�Ƃ��ł���B
�{�����R�n��ŁA�����O����1700m���ō��Ƃ��A���ړ����قƂ�Ǎs��Ȃ��ƌ����Ă���B
�{��͒�R�n�̒J�ԂɌ���ꂽ���z��������x�Ń��J�V�g���{�����l�Ƃɋ߂��n��A���H�����̏ꏊ�Ō����邱�Ƃ������B���̂悤�Ȓn��͂ނ�������Ă���A���J�V�g���{���������n�͌��肳��Ă���ƌ���ׂ��ł���B�{��͉�������Ȃ������ȒJ�Ԃ̎w�W�����Ƃ��ēK���ł���Ǝv����B
����̒�����36�{����苓����ꂽ���A����ȊO�A���������̕W�{�������Ă���̂�37�{���ɕ��z����Ɖ]����B�Y�n�Ƃ��Ă̓��J�V�g���{�ɔ�ׂėy���ɏ��Ȃ����A����̒���������ƁA���ɖ{�B�����Ȗk�ɉ��Ă͖{��̏]�����̐����n�����Ȃ��炸��������A�Y�n�L�^�������������ۂ��������B
�{��͟��o���̂���R����c���̐����n�Ƃ��邽�߁A���H���C�ɂ���ĎY�n�����ł��邨���ꂪ�ɂ߂đ傫���B
�@
�R�j�n�b�`���E�g���{�@Nannophya Pygmaea
���̎�ނ͔����Ŕ������g���{�Ƃ��Ė����ȑO����䂪���Œm���Ă����B���������{���Y��ł͂Ȃ��A����A�W�A�ɍL�����z���A������̓j���[�M�j�A�Ɏ���B���{�͂��̕��z�k���ɂ����邪�A�X����k���Ƃ��A����̒����ł͐�t�E�_�ސ�E�R���E�啪�E�����������41����������ꂽ�B�����������s�ƍ�ʌ����͐�Ŏ�Ƃ��ꂽ�B�{��͔����ŁA�����ɉ��Ĕ������Ղ����߂Ɏw�W�����Ƃ��Ă͓K���ł���B
�{��͂T������W���ɘj��G�߂ɁA�{�B�E�l���E��B�̟��o���̂��鐅�ێ����ɉ��Ă͊e�n�Ŕ�������Ă���A��n��ƌ���ׂ������ł��邪�A�����P�����ō��Y�n�Ƃ��Ċe�n�̎R�Ԃ̎����ɂ����z���Ă���B
�����͐�����قƂ�Ǘ��ꂸ�A�������꒣�������Ď���҂��A������Ԃ͂P���ʁA���͒P�Ƃœ꒣����̐����܂�ɎY������B����܂Ō�����Ă����Y���t�߂ɐÎ~���Ă��邱�Ƃ������B
�c�������̂悤�Ȑ��悩�甭������邪�A�c�����Ԃ͒�n�ł�1.2�N�A���n�ł�2.3�N��v������̂Ƒz�������B
�{��͓��{�̒�n�y�ђ�R�n�̐��ێ����̑�\�I�����ŁA�H���A���Ƌ������邱�Ƃ������A�����̈ꎞ�I�łȂ�����Ȏ��R���̂悢�\���ƂȂ���̂ł���B
����̒����ɉ��Ă͖k�C���E��t�E�_�ސ�E�R���E���m�E�啪�E����̏�������̕������Ă���A�����s�E��ʌ�����ł̈����ɂȂ��Ă���B�]���Ē�n�̎��������݂�������ɂ͖k�C���������đS�{���ɕ��z���Ă������̂ł��낤�B
�{��̐����n�͏�ɒ�n�̎����Ɍ����Ă��邱�Ƃ���A�W�����ӂł͏�ɊJ���Ɠy�n�̑����E�_��P�z�̊댯�ɔ����Ă���B
�{��͐����̈ړ��͎キ�A�S������ʂ��L��Ȑ�������K�v�Ƃ��Ȃ��̂ŁA��r�I���ʐς̕ی�ɂ���Ă���ł͔���������ł��낤�Ǝv����B
�@
�S�j�K���A���V���@NOTOPTERA
�K���A���V�ނ́A�̏W�����̐������Ȃ��A���ɕ��ނɏd�v�Ƃ����Y�̐����������҂̎���ƂɂȂ��Ȃ��W�܂�Ȃ����߁A���ނ����܂�i�W���Ȃ��O���[�v�ł���B���݂܂ł̂Ƃ���A�K���A���VGalloisiana nipponensis�̂ق��A�q���K���A���VG.yuasai�A�I�I�K���A���VG.kiyosawai�A�G�]�K���A���VG.yezoensis�A����э��쌧���ؓ��Y��G.chujoi�i�`���E�W���E���V�Ƃ����Ăі������邪�K���Ǝv���Ȃ��̂ŗp���Ȃ��j�̂T�킪�m���A���茧����͗c���ŋL�ڂ��ꂽ���̂����邪�A���̂Ƃ���^���Ƃ��Ĉ����Ă���B�����ɉ����A�e�n���炱�����Ȃ����L�ڎ�������Ă���B�������Ȃ���A�����̊��m��E���L�ڎ�̎��ʂ́A���Ɍ�������҂łȂ��Ă͂ł��Ȃ��قǍ��G�Ȃ̂ŁA�����ł͓��{�̃K���A���V�ޑS�̂��ЂƂ̎�̂悤�Ȍ`�ɂ��ĂƂ炦�A���������{�����B
���̃K���A���V�ڂ̍����́A���{�̂ق��ɑ嗤���̒��N�������版�C�B�ɂ����ĕ��z���Ă���A����k�A�����J�̖k���R�n�ɂ����z���Ă���Ƃ������قȕ��z�l���������A�܂����̌`�ԓI���ԓI�ȓ_����X���⑶��Q�ƍl�����Ă���B���ہA���̌`�Ԃ͈ꌩ���Č��n�I�ȍ����̂�����v�킹����̂�����B
�K���A���V�ނɂ́A�n�\���̎�Ɠ��A���̎킪����A�O�҂ł͊C�߂����獂�R�ɂ�����܂ł̐X�ђ��̎��������t����y�����邢�͐Ή��ȂǂɎ�Ƃ��Ă��݁A�܂����R�̃K����̊�Ή��A���R�����̕��Ή��Ȃǂɂ��݂���B����A��҂̓��A���̎�́A���A���ł��邱�Ƃ��ς��Ă��鑼�́A�������y�n��Ή��Ȃǂɂ���A�����ꏊ�͍̏������Ă���B�H�ɐl�H������������邱�Ƃ�������̂́A�܂��������R�̂܂܂Ɏc����Ă����n��Ɍ����邱�Ƃ������A���̂悤�ȈӖ��ŁA���R���A�Ƃ��ɍ��R�n�A�R�n�╽�n�̌������сA���R���Ȃǂ̊��̎w�W�Ƃ��Ă͂���߂Ă�����Ă���A�����ɂƂ肠����ꂽ���̂ł���B�������A���ۂɂ͂��̒������̐��������ϊ�ɂ��ɂ������ƂƁA��ʂɂ��̗ނ̒m�������y���Ă��Ȃ��̂ŁA�����ɂ͂�⍢����܂Ƃ��̂��A���_�Ƃ��������悤�B
�K���A���V�ނ́A����������H���ŁA�����悤�Ȋ����ɂ��ރR���V��g�r���V�A�܂��������̗c���Ȃǂ�H�ׂĂ�����̂ƍl������B�����j�ɂ��ẮA���ݒ������ł��邪�A�K���A���V�Ɍ����Ă����A�����琬���ɂȂ�܂łɂ��悻�V�N���O��v������̂Ɛ�������Ă���B
����̂��������́A�k�C���E�{�B�E�l����34���A����ɋ�F����͒��茧���炾�����������B��B�E����n���������āA�̂Ȃ��������͂T���A���Ȃ킿�H�c���E��t���E�_�ސ쌧�E�x�R���E�������ł���B
�K���A���V�ނ̕��ނ̋L�^�́A���܂�ڗ����Ȃ����ł��邱�Ƃ������āA�S���I�ɖR�������A���L�^�̂��̂�������ƁA�O�L�̂̕Ȃ��������̂����A��t�����̂������c�肷�ׂĂ̌��ɕ��z���Ă���A��������B�ł͎������܂łقڑS���I�ɕ��z���Ă���B��������B�̏ꍇ�ɂ͍��ꌧ�̐����ɂ��Ă͊m�F����Ă��Ȃ��B�������t�E���ꗼ���ł����������\���͂���̂ŁA��q���ȓ�̒n�͕ʂƂ��ŁA�K���A���V�ނ͓��{�ɍL�����z���Ă���Ƃ����悤�B�Ȃ��A�̂Ȃ���������������͒�����ЂƂ̎Y�n���m���߂�ꂽ���Ƃ��������Ă����B
�������̌���͂����ނ˗ǍD�ł��邪�A�Ȗ،��̂悤��34�̕��z�n�̂���15�����܂Ŋ����s�ǂɂȂ��Ă���Ƃ��������B���̂ق��X���ł�10�����̕��z�̂����A��������s�ǂ��j��̏ꏊ�ɂȂ��Ă���A�����s��R�����E���R���ł����Ȃ����z����������s�NJ��ɂȂ��Ă��܂��Ă���B
�������̌����́A�X�є��̂ɂ����̂��X���ƈ�錧�������Ă���B�X�є��̂́A�n�ʂ��z�n���A���Ȃ킿���������A���������ꏊ�ɂ͂��̒��͂܂������Z�߂Ȃ��B�܂������̌����Ƃ��Ċό��J�������������̂����Ɖ��R���������A��n�܂��͌��������ɂ����̂��{�錧�������Ă���B������ɂ���y�n���p�̕ω��ɂ́A���̒��͋ɓx�Ɏア�Ƃ����悤�B���̂Ƃ��눫���v���̂������w�I�����ɂ����͕̂���Ă��Ȃ��B
�K���A���ނ̑����́A���������A��������A�������R�������ɕ��z����̂ŁA�n��I�ɂ͂�����x�ی삳��Ă���B
�@
�T�j�^�K���@Lethocerus deyrollei
�����̐����J�����V�Ɠ��l�ɐ����ʼnz�~���A���Ă̍��Y������B100�]��̗��𐅖ʏ�ɒ��o���������̌s�̂܂��ɂ������ĎY�ށB���ʂ̏㏸�ŗ���͐��v���邪�A�ӂ���͐��ʏ�Ɍ����Ă���B�Q�T�Ԍ�śz���A�c�����Ԃ͕s���ł��邪�P�������炢�K�v�ł��낤�B�Ⴂ�c���͐��ʂɏo�Ȃ����A�����͐����Ԃ̐��ʂɕ���ł��邱�Ƃ������B�����͂Ƃ��ɓd���ɔ��鐫���������A�Â�����electric light bug�̖����g���Ă���قǂł���B
�����n�̓��V��K�}�Ȃǂ̔ɖ�������̕����ŁA�����X�V�����r���␅�c�A�܂��͐�̊ɗ����ł���B���̂悤�ȏꏊ�͉a�ƂȂ铮�����L�x�ł���B������x�h�{�^�̐�����D�݁A������n�h�{�^�̐���ɂ͏Z�܂Ȃ����A�܂��a�^�̒r���ɂ������Ȃ��B���ݐ����n���m�F����Ă��鐅��́A�k�͓��{�C���̏H�c�s����A�����m���ł͈�֎s����A��͋{��s�ɓ���͈͂ł���B���̊Ԃɔ����ł��Ȃ��Ȃ��������s�A���I�ɑ��݂���B�����A�V���A�ΐ�A�A����A���Q�A���m�A�����A����̂X���ł���B�m�F����Ă��Ă������n���nj�����Ă��錧�������čl����ƁA�����n���ɓ�k�ɘA��Ȃ�L���я�̖������n�т̂��邱�Ƃ��킩��B�{�B�ɂ�����^�K���̕��z�͂��̒n�т��͂���œ����ɕ������ꂽ��ԂɂȂ��Ă���B�܂��l���̈ꕔ�Ƌ�B�̈ꕔ�ɕ��z��������B�{�B�����n��͓��k�n���̓��암�A�֓��n���ƒ���A�R���̈ꕔ���������͈͂ŁA���{�C���ɂقƂ�Ǖ��z���F�߂��Ȃ��Ȃ��Ă���B�{�B�����n��͖k���̐����A�É����[�Ȑ��̓��C�ƋߋE�n���y�ђ����n�����܂݁A���{�C���ɔ�r�I���z���x���������Ƃ����F�ł���B�l���ł͓��������A��B�ł͍��ꂩ��{��ɂ����Ă̋����я�̕����ɋ͂��ɕ��z���Ă���ɉ߂��Ȃ��B
�����̕s�[���Ȏ��_�ł̌��ʂł��邩��A�����C�����K�v�ł��낤���A�����������Ă���ƕ���Ă��錧�͒���i��ɐ����j�����ŁA����ɂ��ĎR���A��t�A�Q�n�̊e������r�I�����{�킪�������Ă��錧�Ƃ��ċ�������B
�����ɂ킽���Đ����n����r�I�����W���Ă���n��Ƃ��ēȖA���A��������юO�d�A����A�ޗǖk���A���s�A���s�ɋ߂������̑��A���ɂ̂Q�n����l���邱�Ƃ��ł���B����A���A�{��A�H�c�A����A���m�A�����A�L���͐������̒n�_�̑��݂�����Ă���B���k���Ŏ�����̌X�����F�߂��鐶���n������Ă��邱�Ƃ͒��ڂ��ׂ����Ƃ��낤�B
�������̏�ԁ|�ǍD�Ȋ��̂��錧��18�A�s�ǂȊ��̑��݂��錧��26�ŁA�Ƃ������^�K���������ł���Ǝv������̌����錧��34�ł���B
�����n�̑S���܂��͈ꕔ���j��Ă��錧��19���邪�A���͏����Ă��������ׂĂ̐����n���j��Ă���̂��X�A�����A�ΐ�A���Q�A�������A����̂U���ł���B
�������̗v���|����̒����ɂ��A�y�n���p�ɂ�鐶�����̈����܂��͔j��́A���H���݁i�X�P�j��j�A�����E�Z����i���P�j��A�_�ސ�S�����Q�j��A����Q�j��j�A���̑��̓y�n���p�i�Q�n�R�A�x�R�R�j��j�ƂȂ��Ă���A��T�z�ɂ�鐶�����̌������邢�͐�Łi�R��57�A�Ȗ�17�A�����W��ŁA�L���T�A�R�`�S�A����R��ŁA�����R�A�ޗǂQ��ŁA�x�R�P�A����P��Łj����щ͐�̉����ɂ�錸���i�����P�j������Ă���B���̌��ʂ���킩��悤�ɐ��c���̑��ւ̖�T�z���������̌����܂��͐�ł̎���ł��낤�B�����n�̕����I���ł͒����ɐ�łւƘA�Ȃ邪�A�n��I�ɂ͔�r�I���肳��Ă���B���w�����ɂ��ꍇ�͂��̗ʂ���ɂ���ĒZ���ԂŐ�ł��邩�A�������Â��Ȃ����łւ̓������ǂ邩�̂������͂����Ă��A���̉e���͂͂邩�ɍL�͈͂ɂ���ԁB�܂��A���ڐ������̈����Ƃ����Ȃ������m��Ȃ����A���͂Ȑ��ⓔ���ό��n�i���ɎR�n�j�⎩���ԓ��ɐݒu���ꂽ���Ƃɂ��U�E���ꂽ���͂��т����������̂ł������낤�Ǝv����B
�ȏ�̂悤�ɖ{��͂����Ă͑S���I�ɕ��ʂɕ��z�������A�S���̐��c�n�тւ̑��ʂ̔_�^�A�L���ʎ����n�тւ̎E���g�p�̉��d�Ȃǂ�����ƂȂ��āA�S���I�Ȗ��x�ቺ���N���萶���r���ɂ�鉘��������ɒǓ����������A�̐��̌��ŁA�n��I�Ȑ�łɂ�镪�z�n�̕��f��ǒn�����i�݁A���݂ł͖{�B�k���ƒ����n���A�l���A��B�̑啔���Ő�łɋ߂���Ԃ��Ă���̂�����ł��낤�B
�@
�U�j�n���[�~�@Terpnosia vacua
�{��͂S����{���납��A���n���R�т̏��тɏo�����A�����͂T����t�Ŏp���������A�R�n���ł́A�U�����{�܂Ő���������̂�����B�u���[�[���[�[�v�ɋ߂��A�̂�т肵�����Ŗ��A�n���ɂ��u�}�c�[�~�v�A�u�}�c���V�v���ƌĂ����m�̎�ł���B�{�B�A�l���A��B�A�����ɕ��z���A�����̒n���ŕ��ʎ�ł��邪�A�c�����̒������Ɋւ��鎑���͂Ȃ��A���ԓI�m���͔�r�I�R�����B
����̒����ŁA�k�C���Ɖ��ꌧ�ɖ{��̋L�^���Ȃ��̂́A�\�z���ꂽ���R�̌��ʂȂ���A�{�B�ł��A�X�A�H�c�A���A�R�`�A�{��̂T������͕��ꂸ�A���z���Ȃ����A�����Ƃ����̎��_�ł͖������ł���B���݂̖k���͕������ŁA���n�Ȃ�����R�т̃A�J�}�c�тŁA�s�X���̐i��ł��Ȃ��n��ł���A�������F�߂��邪�A���Ȃ��ƕ���Ă���B����̎��������̕ł́A��B�{�����ł́A�N���}�c�A�A�J�}�c�A�����E�L���E�}�c�̗тɔ�r�I���ʂŁA�}�c�тւ̔_��̋U�z���A�{��ɂǂ̂悤�ȉe����^���Ă��邩�͕s���Ƃ��Ă���B�{�킪���v���܂œ쉺���Ă���\�����������A���݁A���v���ɎY����L�^�͂Ȃ��B
�{��̐����ƔɐB�̂��߂ɂ́A���i�A�J�}�c�A�N���}�c�A�����E�L���E�}�c�j�̎��R�т��K�v�ŁA�s�s�����i�ނƋ��ɐ����n�����߂��A�u���������v�ɂ�鏼�͂�̌�����n���ł́A���̊g��Ƌ��ɁA�}���Ɍ̐����������A���łɂ��̖������S���₦���n�������Ȃ��Ȃ��B
�@
�V�j�M�t�`���E�@Luehdorfia japonica
�N�P��̔����A�����̏o�����͊֓��Ȑ��̒g�n�ł͂R�����{����S���A���z�̖k���ɋ߂��n���W���̍����ꏊ�ł͂S�����{����T���ɂ����Ĕ�������B�c���̐H���͂��ꂼ��̔����n�Ɏ�������J���A�I�C�ނł��邪�A����n��ɕ�����̃J���A�I�C������ꍇ�ɂ́A���ꂼ��̎�ɂ��đI�D���̈Ⴂ�����邱�Ƃ��m���Ă���B�܂��n��ɂ���Ă̓E�X�o�T�C�V����H���Ƃ���B�����̋z���A���̓J�^�N���A�X�~���ށA�T�N���ށA�V���E�W���E�o�J�}�A�{�P�A�I�I�C���J�K�~�A���L�c�o�L�ȂǁB���͐H���̐V�藠�ʂɐ��Ȃ����\���������߂ĎY�݂����A�z�������c���͎�߂̊Ԃ͗t���ɏW�����A�ېH���W�c���Ȃ��čs�Ȃ����A���߈ȍ~�͒P�Ɛ��������s�Ȃ��ӏt��匉��A�āA�H�A�~��匑Ԃʼn߂����A�����t�ɉH���o���̌o�߂��Ƃ�B
�M�t�`���E�̎�Ȕ����n�͋u�˒n�`��R�n��̃R�i���E�N���E�N�k�M�Ȃǂ̒g�ѐ����t�L�t���тƂ��̎��ӂŁA���t�ɗz�����я��ɒB����悤�Ȗ��邢�G�ؗтł���B�n��ɂ���Ă͏Ɨt���ѓ����邢�͂��̎��ӂ̗z�n�A���邢�͉��ѐ��̗��t�L�t���тɂ���������B�����I���ɂ͖{��͔������Ȃ��B
���k�n���̑����m���n��ɂ͖{��͂܂��������z���Ȃ����A���{�C���ł͐V���E�R�`������C�ݒn�тɉ����Ėk�サ�A�H�c���œ�[�̗R���C�ݓ암�ɒB���A���̒n���{��̕��z�k���ƂȂ�B�֓��n���ŃM�t�`���E�����z����̂͐_�ސ쌧����ѓ����s�����n��݂̂ŁA���̌����ɂ͂܂��������z���Ȃ��B�����s�����q�s�����R���瑽���u�˂ɂ����Ă̈�т͂��Ă͖{��̎Y�n�Ƃ��Ē����ł��������A���݂ł͔����q�s���[���̗ыƎ�����������тɋ͂��ɑ��݂��Ă���ق��A��L�̑S�n�悩��قڊ��S�ɐ�ł����B���̌����͐����n�̑�n���ƊJ���ɂ����̈����ł���A�Ƃ��ɍ����R�ł͋ƎҁE�}�j�A�ɂ�闐�l�̉e�����傫���ƌ����Ă���B�x�R�E�ΐ�E����̏������ł͋u�˒n�`��R�n�̃R�i���̓т���Y�n�Ƃ��ĕ��z�͍L���̐��������A���ݓ��{�ł����Ƃ��L�x�ɃM�t�`���E��������n��̈�ł���B
�ߋE�E�����n���ł́A�ߍ]�~�n���ӂ̎R�[�E�R�n�тƂ��ɗ鎭�R���R�[�A����E���������A�u�˒n�`��R�n�Ȃǂ̖L�x�ȎY�n�����݂��邪�A��s�s�̎��ӂ𒆐S�ɒ����������������n����ł����n����������݂���B���̌����Ƃ��ẮA��Ƃ��Ď��R���̔j��Ɨ��l�ɂ���Ƃ����Ă���B
�Ȃ��A�R�����̎R���s�Ɣ��s�����Ԑ����{��̕��z�̐����ƂȂ�B�l���E��B�n���̌Â�����ɗ��n�悩���̕s�m���ȋL�^�����邪�A���̌�̖Ȗ��Ȓ����ɂ���Ă��{��͂܂�������������Ȃ��̂ŁA�{��͂��̒n��ɕ��z���Ȃ��ƍl������B
�M�t�`���E�̎�ȎY�n�́A�u�˒n�`��R�n�̑��t�ɗz�����я��ɂ������ނ悤�Ȗ��邢���t���̎G�ؗтŁA�R�i���E�N���E�N�k�M�Ȃǂ̎������Ƃ���т������B�G�ؗт����̂��ꂽ��ɐA�͂��ꂽ�X�M�E�q�m�L�̗c�ߗтɂ��ɂ������������邪�A����炪���炵�ėѓ��ɗz���������ʂ悤�ɂȂ�Ɩ{��̔����͏I������B���������ēK���ɎG�ؗт�����A�G��ɂ��т̌`�����u���I�ɍs����悤�Ȓn�悪�{��̐����n�Ƃ��Ă͂����Ƃ��K���ł���A�̐��������B�ߔN�A�d�Y�ނ̎��v�̌����ɂ��o�ω��l�̂Ȃ��Ȃ����G�ؗт��X�M�E�q�m�L�̐A�ђn�ɂ������邱�Ƃ��������ƁA�S���t��A�Z��n�����Ȃǂɂ�鎩�R�J���A����Ƀ}�j�A�ɂ��H���̌@���A���E�����̗��l�ɂ���Ė{�킪��ł܂��͒������̐����������n��͂���߂đ���
�@
�W�j�q���M�t�`���E�@Luehdorfia puziloi
�����͔N�P��̔����A�{�B�ł͂S�`�T���A�k�C���ł͂T�`�U���ɏo������B�c���̐H���͖{�B�ł̓E�X�o�T�C�V���A�k�C���ł̓I�N�G�]�T�C�V���B�����̋z���A���̓J�^�N���A�X�~���ށA�T�N���ށA�L�N�U�L�C�`�����\�E�A���}�G���S�T�N�ȂǁB�����j�̑�v�͑O��M�t�`���E�Ƃقړ������B
�����n�̊��͉��ѐ��Ȃ��������ѐ��̗��t�L�t���сi�܂��͐j艍���сj�A�J���}�c�A�ђn����т��̎��ӂŁA�����̏o�����ɂ͗ѓ������邭�A�z�����я��ɓ͂��悤�ȏꏊ�ł���B�M�t�`���E�͒g�ѐ��̗��t�L�t���сi�G�ؗсj����Ȑ������Ƃ�����̂ŁA�ʏ헼��͖��Ăɂ��ݕ����Ă��邪�A�M�t�`���E�̕��z�����їщ����ɂ܂ł̂ڂ�n��i����E�R���E�V���E�����E�R�`���̈ꕔ�j�ł͋ǒn�I�ȗ���̍����n���m���Ă���B
�k�C���A���k�n���A�{�B�����ɕ��z�A��ʂɌ̐��̑������̂ł͂Ȃ����A�D�K�Ȋ��̎Y�n�ł͖L�Y����Ƃ��������B
�H���̗L���͖{��̕��z���x�z����ő�̗v���ł��邪�A�H���������Ă����z���Ȃ��n��͑����A�q���M�t�`���E�̕��z�ɑ��̏������傫���֗^���Ă��邱�Ƃ͋^�����Ȃ��B���т̔��̐Ւn�ɂ͈ꎞ�I�ɐH���̃E�X�o�T�C�V���A���������ƂȂ�J�^�N�������͂𑝂��A����ɂ�Ė{������̌̐��𑝂��̂���ʂł��邪�A���̐Ւn�ɃX�M�̐A�т��s���A���ꂪ�������ďt�ɂȂ��Ă��я��ɗz���������܂ʂ悤�ȏ�ԂɂȂ�Ɩ{��͏��ł���B�X�M�E�q�m�L�̑��сA�y�n�J���Ȃǂ̊������ɂ���ŁA�������͒������̐����������Y�n�͏����Ȃ��B�܂��}�j�A�ɂ��H���̖x���A���̑�ʍ̎�ɂ���Č̐��̌���������Ă���n��������B
�@
�X�j�I�I�����T�L�@Sasakia charonda
�����͔N�P��̔����A�g�n�ł͂U�����{������A����n�ł͂V�����{������o������B�֓��ȓ�̒g�n�ł͂܂�ɏH�X�`10���ɕ����I�ȑ�Q���̔������邱�Ƃ�����B�c���̐H���͒g�n�ŃG�m�L�A����n�ł̓G�]�G�m�L�B���͂V�`�W���ɐH���̗t�A���}�Ȃǂɐ��݂����A�U�`�V���ŗc�����z������B�c���͏H�܂łɒg�n�łS�߁A����n�łR�߂ɒB���A�H���̗t�����݁A������悤�ɂȂ�ƁA�̐F�͗ΐF���犌�F�ɕς�A���}���犲�������Ēn��ɂ���A�H�����ۂ̗��t���ʂɐÎ~���ĉz�~�ɓ���B���t�H���̉藧���ƂƂ��ɓ~����肳�߂��c���͐H���ɂ̂ڂ�ېH���J�n���邪�A�z�~��ŏ��̒E��ɂ��ΐF�ƂȂ�A����ɂ�����x�E�炵�ďI�ߗc���ƂȂ�B匉��͒g�n�łU���A���n�łV���ɍs�Ȃ��A15�����O��匊��̌�A�H�����Đ����ƂȂ�B�����̓N�k�M�E�R�i���E�J�V���E���i�M�ށE�^�u�Ȃǂ̎��t�ɗ��W�A���邢�͕��ʁA�����̕��E���̂ɂ��W�܂�B���A�Ԃɂ͂��Ȃ��B
�{��̕��z�̒��S�͉��с`�g�т̃R�i���E�N�k�M�E�N���E�~�Y�i���Ȃǂ́A���邢�͂����̎�����܂ގG�ؗтł���B�������c���̐H���i�G�m�L�E�G�]�G�m�L�j�̑��݂͕K�{�����ł���B������{�ł͏Ɨt���тɂ������邱�Ƃ����邪�A���������ꏊ�ł͌̐��͏��Ȃ��B
�k�C���̈ꕔ�A�X������B���������k���ɂ����ĕ��z�A���x�A�B��E�Γ��E�ɓ������Ȃǂ̗����A���v���ȓ�̓쐼�����ɂ͕��z���Ȃ��B�n���ʂ̐����͎��̂Ƃ���ł���B
�k�C���n���G�k�C���Ő����n�Ƃ��Ēm���Ă���͎̂D�y�s�Ɨ[���s�̎��Ӓn��A�Ŗk�̎Y�n�͐Ύ�x���l�v�����c�ł���B
���k�n���G�S�����ɕ��z���邪�A��ʂɎY�n�͓_�݂��A�̐����Ƃ��ɑ����Ȃ��B���n�`��R�n�̎G�ؗт���Ȑ����n�ł��邪�A�����̎G�ؗт͐A�сA�J���ɂ���Ĕj��邱�Ƃ������A�قƂ�ǂ̒n��Ō̐��̌���������Ă���B
�֓��n���G�S�s�����ɕ��z���m���Ă��邪�A��ʂɕ��n�`��R�n��̎G�ؗтɑ����A1000m�������悤�ȍ��W���n�ɂ͐������Ȃ��B���n�`�u�˒n�̎G�ؗт͊J���ɂ�莟�X�ɔj��Ă���̂ŁA���݂ł͑����̐����n�Ő�ł����B�����E��ʂł͕�����̎G�ؗтɑ����������A�������s���ł͊��S�ɐ�ŁA�����̔����q�s�A�~�s�A�������S�ɐh�����Ĕ����n���c����x�ɂ܂ŏ͈������Ă���B�_�ސ쌧���ł������ȓ�̕��암����قƂ�ǂ��̎p���������B
�����n���G�S�����ɕ��z���m���邪�A���n�`��R�n��̎G�ؗт̔��̂��e�n�Ői�s���A�����̒n��ł͐�ł܂��͒������̐������������B�������R����������ł͂Ȃ������̑��Y�n�i�Ⴆ�Γ��t��j���c���Ă���A���ݓ��{�ł����Ƃ��̐��̑����n��̈�ł���Ǝv����B
�ߋE�n���G�S�{�����ɕ��z���m���邪�A�s�s���ӂ̐����n�ł͊J���ɂ���Ďp�����������������B��ʂɌ̐��͑����Ȃ��B�ޗǁE�a�̎R���ł͎Y�n�͏��Ȃ��A���Ɍ���ˎs��ʁA�쐼�s�A�_�ˎs�k��Ȃǂł͐�ŁA�܂��͂قƂ�ǎp���������B
�����n���G�S�����ɕ��z���m���邪�A���R�����ł͐��암�ɂ͐������Ȃ��B���n���Ɠ��l�ɕ��n�`��R�n�̎G�ؗт̔��̂ɂ���Č̐��̌����������ł��邪�A�s�s���痣�ꂽ�R�[���̎G�ؗтɂ͂Ȃ������̔����n���c���Ă���B
�l���n���G�S�����ɕ��z���m���邪�A���n�`�u�˒n�ɂ͂قƂ�Ǖ��z�����A�R�n�E�R�Ԃ̎G�ؗтɐ������c���Ă���B�S�ʓI�Ɍ̐��͏��Ȃ��B
��B�n���G��B�n���ł͕����E�F�{�E�啪�E�{��E�����������ɎY�n���m���邪�A����E���茧���ɕ��z���Ȃ��B��B�n���ł͕��n�`�u�˒n�ɔ������鏊�͂Ȃ��A������̌����ł��R�ԕ��ɂ��̐����͌�����B�̐��͑����Ȃ��B�{�茧���юs�A�{�ؑ��A������������s�A�o���s�����̕��z�̓���A�����R�[����т�����ȓ�̒n��ɂ͕��z���Ȃ��B
���k�n�����֓��n���ɂ����ẮA�I�I�����T�L�͐l���߂��̕��n�`�u�˒n�̃R�i���A�N���A�N�k�M�A�A�x�}�L�Ȃǂ̎G�ؗт���Ȑ����n�Ƃ�����̂ŁA�܂���荂�n�̃~�Y�i���Ȃǂ̎G�ؗтɂ����̐������͋y��ł���B���n�`�u�˒n�̎G�ؗт͐d�Y�ނ̎��v�̌��ނ���o�ω��l�̒Ⴂ���̂ƂȂ�A���̂��ߔ��̂���ăX�M�A�q�m�L�̐A�т��i�݁A�܂��Z��p�n�Ƃ��ĊJ������A�I�I�����T�L�̐����ɍD�K�Ȋ��͑�Ђɔj��A�܂��͉��ς��i�s���Ă���B���������đ����̒n��Ŗ{��͐�ł��A�c�����R�n�ł����������̌̐��͌��������B���{�̐��암�A�Ƃ��Ɏl�����B�ł͖{��̎�Ȑ����n�͎R�Ԃ̎G�ؗтł��邪�A�����̏ꏊ���X�т̔��̂��Ђǂ��A���̌̐��̌����͒������B
�@
10�j�Q���W�{�^���@Luciola cruciata
�n��ɂ��A�N�o�ߐ��͑����قȂ�悤�ł͂��邪�A�����̊������Ԃ͂����ނ˂U����{����V�����{�ɋy�сi��F�ł͂T����{�A���k�n���k���ł͂V����{���犈�����ɓ���j�A�w�C�P�{�^���̊������ɂ���ׂ�ƁA�S�̓I�R�T�ԂقǑ����B
���͐��ӂ̃R�P�Ȃǂɒ��a0.5mm���炢�̉��F�������݂���B�Y�����͕���500���炢�Ƃ�800�`1200�Ƃ�������B�n��I�ȍ�������̂�������Ȃ��B��P�����i26�`30���j�łӉ������c���͗����ɓ���A�J���j�i���a�ɐ������͂��߂�B�~���z���A�R�A�S�����܂łɂU��E�炵�A�[���ɐ��������c���͉J�̓���I��ŏ㗤�A�y���ɐ�������匉��A��30����匊����o�ĉH������B�H���͗Y��������P�T�ԑ����B�H�����������͎��Y�Ƃ������̂ނ����ʼna�͂Ƃ�Ȃ��B��������10�`20���ԂŎ������I����B
�{��̐������Ƃ��ẮA�����̑��݂��邱�ƁA�a�ƂȂ�J���j�i���������邱�ƁA���ӂɂ�匉��ɓK����n�ʂ�����A�Y���ɓK�����R�P�����炵�Ă��邱�ƂȂǁA�����Ƃ��O�̏������K�v�ł���B�܂����Ɋւ��ẮA�N�Ԃ�ʂ��Đ������T�`21���͈̔͂ɂ���APH6.5�`7.8�A�����̔ɖ��Ă��Ȃ����ƂȂǂ������ƂȂ�B
�Q���W�{�^���͖{�B�E�l���E��B�A����ɗ������ł͈����E���x�E�B��E�Δn�E�W�H���E�������ȂǂɍL�����z���邪�A�k�C���A�ɓ������A���A�ܓ�����ю�q���ȓ�̓쐼�����ɂ͌����Ȃ��B
�����n�Ƃ��ĕ��ꂽ�n�_�́A���Ɍ��ȊO�͑S�ʓI�ɏ��Ȃ����A����͏���R�����������ƂɌ���������炵���B���ۂɂ́A����ɑ����̐��ݐ����n�����݂��Ă���ƍl���Ă悢�Ǝv���B
�����n�̏����E�̐������������炵���v���Ƃ��ẮA�_��g�p�i�R�`���ق�17���j�A�C���i���Ȃǂ̂��߂̓ŗ����i�X�j�A�E�L�܂ɂ��~���C���K�C�쏜�i�R���j�A�q��E�{�؏�Ȃǂ���̉��������i�X�E���j�ƒ�r�������i���ȂǂR���j�i���̂ق��A�P�ɐ������Ƃ������̂������ȂǂU������j�A�ӐA�y�؍H���ɂ��y�������i�X�ȂǂR���j�A��n�����ɂ�闬���̏����E�y�������i��t���ȂǂR���j�A�썻���̎�i�����j�A�͐�E�p���H�̉��C�i�������Ȃ�18���j�A���l�i�R���ȂǂS���j�A���Q�i����j�Ȃǂ��w�K���ꂽ�B�����ɋ�����ꂽ�v���́A�e�n�ɂ�����Q���W�{�^���̌����ƁA�����ꏭ�Ȃ���W��L���Ă�����̂ƍl������B
�����[���̂́A���h�_����_���̌��݂ʼn͐�̐��ʂ����肵����A�쏰�����肵���肷��ƁA���ꂪ�Q���W�{�^�������ɍD���ʂ������炷�Ƃ���݂̂�ꂽ���Ƃł���i�a�̎R�A���Ɂj�B
���ʓI�ȕ����͏��a30�N��ɐ����n�̏��ł�z�^���̌̐��̌����������A�ŋ߁A������������Ƃ������Ƃł���B���̗��R�Ƃ��āA���͂Ȕ_��̎g�p�Ƃ̊W���w�K����Ă���B
�Q���W�{�^���͌Â�������{�l�ɐe���܂�Ă������{�̑�\�I�����̈�ł���B�R�ԕ��݂̂Ȃ炸��n�ł��A�����ɐ���������Q���W�{�^���͐������Ă����B�]�ˎ���̋L�^������ƁA������23����ɂ��z�^���̖������U������A���̏����̂Ԃ��Ƃ��ł���B�������A���݂ł͓s�s�悩��͂قƂ�ǎp�������A�܂��c���n�т�R�ԕ��ł���ł��邢�͌��������n�����Ȃ��Ȃ��B
�����������ނւ̑Ή���̈�Ƃ��āA�Q���W�{�^���̗{�B���͂��߂�ꂽ�B�{�B�͏��a12�N�A��R�s�̓��s�Y���ɂ���Ă͂��߂Ď肪����ꂽ���A���a40�N��ɓ����Ă���͑S���I�ɂ��̎��Ƃ����y���͂��߂��B�n�������̂̎��R�ی슈���Ƃ��āA���E���E��w�Ȃǂ̃N���u�����̈�Ƃ��āA���邢�͂܂��A�u����v�A�u�����v�A�u�ۑ���v�A�u������v�Ȃǖ��Ԓc�̂̊����Ƃ��āA�ډ��}���ɐi�W���Ă���B�������A���̂悤�ȗ{�B�A�����ɂ͖�肪���Ȃ��Ȃ��B
���Ȃ킿�A�̐������������n��ŁA�n��̃z�^����{�B���ėc�����������̂ł���Ƃ������A���łɐ�ł����n��܂��͌��������n��ɁA���̒n�悩��z�^�������ĕ��������ꍇ�A�`�Ԃ����łȂ��A�����炭���ԓI�ɂ���l�̂��̂ł͂Ȃ��A���{�̃Q���W�{�^���̎��R���z���h�����錋�ʂɂȂ邩��ł���B���݁A���łɗ��Y���đS���e�n�ɂ��������Ǝ҂��猻���Ă���B����́u�z�^���̉ƒ{���v�ȊO�̉����̂ł��Ȃ��B
���ꂼ��̒n��ɁA���ꂼ�ꐶ�������Ă������j�I�ȃz�^���̐���������Ă����A�͂��߂Ď��R���A�����ăz�^������������ƂɂȂ�̂ł���B
�@
�R�D�w�W�����ނ̕��z�E�������̏�Ԃ���݂��s���{���̊��f�f�̎���
�@�w�W�����ނ̐������́A�R�n���畽�n�Ɏ���܂ł̉�������ς��i��ł��Ȃ������i�k���A����A���n�A�r���j�⎩�R�т�тȂǂŁA�����͐l�Ԃ̐�����̒��ɓ邶�ݐ[�����R���A����Η��R�E�c���I���R�̑�\�I�v�f�Ƃ݂Ȃ�������̂ł���B
�@���̂悤�Ȋ��͔䗦�͈قȂ��Ă�������̓s�{���ɂ����݂���ƍl�����A�w�W�����ނ��قڑS���ɕ��z������̂ł��邩��A���������̍��������ׂāA��r�I�e�ՂɌ��o�����Ƃ��ł���Ȃ�A���̒n��͗��R�E�c���I���R���ǍD�ȏ�Ԃő��݂��Ă���ƍl���邱�Ƃ��\�ł��낤�B�t�ɖ{�����Y�n��ɐ�������͂��́A�����̎w�W�����̐������m�F�ł��Ȃ�������A�ɒ[�ɏ��Ȃ��ꍇ�́A�Ή�������������Ȃ����j��Ă��邩�A�������㑶�݂��Ă��Ă����Y��̐����������Ȃ����I�ω��i�����j�������Ă�����̂Ƃ݂Ȃ����Ƃ��ł��邾�낤�B
�@���̂悤�Ȋϓ_����A�s���{�����ƂɁA����ݐ[�����R�����o�����X�̂Ƃꂽ��Ԃɂ��邩�ǂ������A�����̐�����Ԃ�ʂ��Đf�f���邱�Ƃ����݂��B
�@�A���A10��̎w�W�����ނ̂����A�K���A���V�ڂ͓���ݐ[�����R���̎w�W�Ƃ��Ă͂��K�i����������A���t����y���A�Ή��Ȃǂ��ꏊ�Ƃ��邽�߁A����߂Ėڂɂ��ɂ������ƁA��ʂɂ��̗ނɊւ���m�������y���Ă��Ȃ����Ɠ��̗��R�ŏ\���ȏ�����Ȃ������Ǝv����̂ŁA���͂̑Ώۂ���͏��O�����B
�s���{�����ɓ���݈Ղ����R�����A���a�̂Ƃꂽ��Ԃő��݂��Ă��邩�ǂ����𐄒肷�邽�߁A���̂悤�Ȋ����ǂ̒��x���݂��邩�̎w�W�Ƃ��āA�����̐����n�̋K�͂��A�����Ă��ꂪ�ǍD�ȏ�Ԃŕۂ���Ă��邩�ǂ����̎w�W�Ƃ��āA�����̐������̏�Ԃ����ʂ��W�v�����B
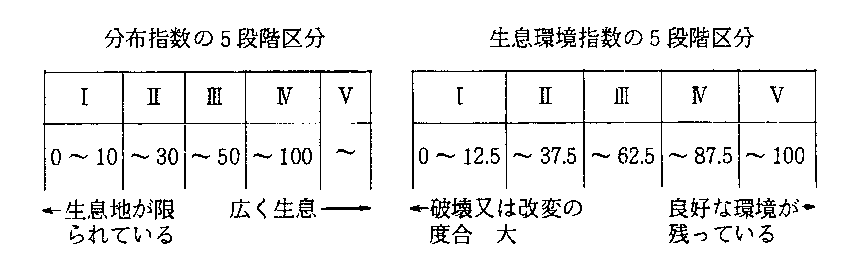
�P�@�����n�̋K�͂̔c���i���z�w���̎Z�o�j
�w�W�����ނ̐�����������́A������̓s���{���ɂ����݂�����Əq�ׂ������̑��ݔ䗦�͂����炭���܂��܂ł���A����Ɏ��ۂ̐����n�ƂȂ�A�펩�̂̒n���I���z�����ɑ傫���e������B�w�W�����͑S���I�ɕ��z������̂���I�ꂽ���A�k�C�����牫��܂ŕ��z����̂̓^�K���݂̂ł������i������������{��A�lj����ɂ���Ėk�C���ɂ����鐶�����m�F���ꂽ���̂ł���j�B
�}�Q�|�V�|11�͎w�W�����ނX��̕��z��s���{���ʂɕ\�킵�����̂ł���B���̐}���番��Ƃ���A�Ώێ�X��͖{�B�A���ɒ������𒆐S�ɕ��z���Ă���A��ސ�����݂ď\���ȕ��͂��\�Ȃ͖̂{�B�݂̂ŁA�k�C���A����͂��̕��͑Ώۂ��珜�O������Ȃ��B���{�B�̈ꕔ�A�l���A��B�͏��̌��@�ɗ��ӂ��āA���͌��ʂ����߂���K�v������B
�]���ē��k�n����l���A��B�͊���̎�̕��z���E�n��ƂȂ邪�A�����̒n��ɂ����Ă͈�ʂɓ��Y��̕��z��͋����̂ŁA�����n�̋������璼���ɓ���ݐ[���������Ȃ��Ɣ��肷�邱�Ƃ͂ł��Ȃ��B
�s���{���Ԃ̔�r���s���ۂɖ��ƂȂ�̂́A�����n��̍L����̕\�킵�����\�����ꂳ��Ă��Ȃ����Ƃł���B�W�v�i�K�ł̓��b�V���ւ̕ϊ����s��ꂽ���A�������_�ł́A�����n��̕\���͊��m�̖ʐςɉ������̊O�����͂ނ��i������j�A�_�\�����̕��@���Ƃ�ꂽ�B�O�҂̏ꍇ�A�L������ߑ�ɕ\���������ł����A�s�m���ȏ���̌`�ŕ\������邱�Ƃ��܂܂���B����A�_�\���͋t�ɍL������ߏ��]������X��������A���̕��@���Ƃ����ꍇ�A�s�m���ȏ���O����Ղ��B
���̂悤�ȕ\�����@�ɂ�����ɗ͎揜�����߁A�s���{�����̐����n�̉ӏ����ƁA�����n�̍L��������b�V���ɕϊ������ۂ̃��b�V�����Ƃ̑���i�j���ς����߁A��������ꂼ��̎�̓��Y�n��ɂ����镪�z��̋K�͂������w���i���z�w���kD.I�l�j�Ƃ����i�t�\�Q�A�}�Q�|�V�|12�j�B����ɂ͔j�ꂽ�����n���J�E���g����Ă���̂ŁA������x�ߋ��ɑk�������_�ł̐����i�����_�Œm�肤�����̌����z�j��\�킵�Ă���B
�Q�@�������̏�Ԃ̔c���i���w���̎Z�o�j
�������̏�Ԃɂ��Ắu�������̌���v�Ƃ��āA�����n���ƂɁu�ǍD�v
�u�s�ǁv�u�j��v�̔��肪�����҂ɂ���ĂȂ���Ă���B�s���{�����Ƃ̐������̏�Ԃ𐄒肷��̂ɁA����c���̂Ȃ��ꂽ�����n�i�u�s���v�����������́j�̉ӏ����A���b�V�����ɑ���u�j��v�y�сu�s�ǁv�ȏ�Ԃɂ��鐶���n�̉ӏ����A���b�V�����̔�����߁A�P�Ɠ������R�ɂ�葊��i�j���ς��Z�o���A��������w���iE.I�j�Ƃ����i�t�\�Q�A�}�Q�|�V�|12�j�B
�A���A���̕��@�ŋ��߂��l�͈����Ȃ����͉��ς̒��x��\�����̂ł���̂ŁA�ǍD�ȏ�Ԃ�\���ꍇ�́A���̒l��100���猸�������̂�p�����B
���̏ꍇ�ɂ��P�Ō��������悤�ɁA�����̒n���I���z�����ɗR������n�捷���l�����Ȃ���Ȃ�Ȃ��B���Ȃ킿�A���z���E�n��ł̐��������͉������E�ɋ߂����Ƃ������A���̂悤�ȏꍇ�ɂ́A���z�̒��S��i�e��̐����������œK�͈͂ɂ��邱�Ƃ������j�ł́A�قƂ�lje���̂Ȃ��悤�Ȋ��̕ω����A�v���I�ȑŌ��ƂȂ邱�Ƃ�����A���z���E�n��̊��w���͂�����x�������čl����K�v�����邾�낤�B
�R�@�s���{���ʃ��[�_�[�`���[�g�̍쐬
�w�W�����ނ̕��z�A�������̏ɂ���ĕ\�킳���g�|�^���Ȏ��R���̏�s���{���P�ʂŔc�����邽�߁A�P�Q�ŎZ�o�����w����p���ă��[�_�[�`���[�g���쐬�����B
�쐬�ɓ������ẮA�u���z�w���v�A�u���w���v�����̂Ƃ���敪���A���̃����N�ɉ����āA�Y������햼�̈ʒu�Ƀv���b�g�����B
|
�e�s���{���̃��[�_�[�`���|�m�y�ѕ\�����e�̏ڍׂ��}�Q�|�V�|13�`14�Ɏ����Ƃ���ł���B
�i�Q�j���͌��ʁi�w�W�����ނ̕��z�A�������ɂ��s���{���̊��f�f�j
���f�f������ɓ������Ă͍����ނ̐����n�̑召�A�������̗ǁA�s�ǂ肷�����K�v�ł���B
���݂̂Ƃ��낱����w�I�ɋ敪���邽�߂̎�i�A�m���͑��݂��Ȃ��̂ŁA����I�Ȃ��̂ɂȂ炴��Ȃ��B�����ł́A�ꉞ���z�w���̏ꍇ�͇U�������A���w���̏ꍇ�͇U�ȉ������ꂼ�ꐶ���n�������Ă����ԁA�������̉��ςȂ����j�i�s���Ă����ԂƂ݂Ȃ��ċ敪���s�����B
�Ȃ��\�Q�|�V�|�S�͊e�����N�ɑ�����s���{�����ł���A�^�K���̕��z�w���A���w���̒Ⴂ���̑����̂��ڗ��B
�쐬���ꂽ���[�_�[�`���[�g����A�\�Q�|�V�|�T�Ɏ����ޕʂ��Ȃ��ꂽ�B����ɂ��ƁA�S��̐��������ǍD�ȏ�Ԃɂ���̂͊��A�O�d�A���s�A���R�A�L���A�R���̕{���ŁA�����̒n��ł́A����ݐ[�����R�������a�̂Ƃꂽ��Ԃő��݂��Ă���Ƃ�����B�A���\���Ȍ��n�m�F���s��ꂸ�A�啔���̐����n���A�������Ɋւ��s���Ƃ��ċL�^����Ă���ꍇ�́A���w�������Ԃf���Ă��Ȃ��ꍇ������B���̂悤�ȏꍇ�ɂ́A���������Q�l�ɂ��ĕ���s�����B
�܂��A������̎w�W��̑S�����͈ꕔ�����R���z���Ȃ��n�悪���邪�A���̏ꍇ���̎w�W�����̂������ɏ]�����B
���̂悤�ȑ���ɂ��A����敪�ɒlj����ꂽ�n�悪�i�@�j���Ɏ�����Ă���B��L�̕{���������s���{���ɂ́A�X��̎w�W�����ނ̕\�킷����ݐ[�����R���̂����ꂩ�̕����ɁA�j��∫���̒����݂���ƍl������B�\�Q�|�V�|�T�ɂ̓g�[�^���Ȋ��̑��ɁA�W�敪��݂��A���ꂼ��̋敪�ɑ���������ǍD�ȏ�Ԃɕۂ���Ă���A���邢�͉��ψ������i��ł���Ǝv����s�{�����������B
���𐅕ӂƗђn�ɕ������ꍇ�A���ӊ����ǍD�ȏ�Ԃɂ���̂́A���m���P���݂̂ł���̂ɑ��A�ђn���ǂ���Ԃɂ���̂́A�x�R�����͂��ߑ�������A�ΏƓI�ł���B�n�`�I�ɂ݂ĎR�n�`��R�n�̊����ǍD�Ȃ̂́A���Ƌ{��ł���B�������{��̓��J�V�g���{�����R���z���邩�ۂ����s�m���ł���̂Ŏ�̋^�₪�c��B��R�n���畽�n�ɂ����ėǍD�Ȃ͕̂������݂̂ł���B
�R�n�����R�n�ɂ����Ă̐��ӊ����ǍD�Ȃ̂́A��ʁA�V���A�É��A�a�̎R�A�R�`�A�{��̂V���ł���A���̋敪�͗ђn�����ǍD�ȋ敪�Ɏ����ő����B�ђn�������ǍD�Ȃ̂͊��ł������B��R�n���畽�n�ɂ����Ă̐��ӊ����ǍD�Ȃ̂͑��A����A�����ăn�b�`���E�g���{�����R���z���Ȃ���Α啪�̂P�{�Q���ŁA�ђn�������ǍD�Ȃ͓̂ȖA�R���ł������B
�@
�S�D�w�W�����ނ̐������̔j��E���ϗv����̌Q�̐�ŁE�����v��
�@�����[�̋L�q����A�������̔j��E���ς̗v����̌Q�̐�ŁE�����̗v����ǂݎ��ތ^�����A�u�j��v�A�u�s�ǁv�̕\���̂��鐶���n�ƑΉ������A�s���{�����ƂɃ��b�V�������W�v�����i�����ҁj�B
�@�����[�̋L�q�́A�X�̐����n�ƑΉ����Ă��Ȃ������A���łɏq�ׂ��悤�ɕ��z�}��̐����n�̕\�����@�͈�肵�Ă��Ȃ��̂ŁA���̏W�v�l�������āA���Y��ɑ���e��v���̉e���̒��x���ʓI�ɔc������͍̂���ł���B�����ŁA�w�W�����ނ̊����̎�ނƂ��̒��x���A���ΓI�ɔc�����邽�߁A�v�����Ƃɐ������b�V�������ɑ���S����ƁA�v���̎w�E���ꂽ�s���{�����́A��������s���{�����ɑ���S��������߂������ŁA�����̑���i�j���ς����߂��i�\�Q�|�V�|�U�j�B
�@����ɂ��ƁA���������̂��̂�j��v���i�����I���ρj�Ƃ��ẮA���H���݁A�X�є��́i�A�т��܂ށj�A������n�����A�͐���C�i�_�����݂��܂ށj����Ȃ��̂ŁA�������̎�������������v���i���w�I���ρj�Ƃ��ẮA��U�z������p���E�H�Ɣp�����ɂ��͐�̉�������������B�Ȃ��A�����I���ς̈�v���Ƃ��āA�ό��J�����������Ă��邪�A����́A��̓I�ɂ͘H���݁A�X�є��́A������n�������̉��ύs�ׂ��܂ނ��̂Ǝv����B
�@�����̗v���ɂ���ĉe������p�x�́A��ɂ���ĈقȂ�A���J�V�g���{�A���J�V�����}�͓��H���݁A�X�є��̂���ł���A�Q���a�{�^���͉͐���C�A��U�z�ɁA�n�b�`���E�g���{�͌����E��n�����i�ɂ�鎼�n�̖����j�ɁA�^�K���͖�U�z�ɋ����e�����Ă���B
�@�ђn�����Ƃ�����̂́A�����ނːX�є��̂�A�����E��n�����ɂ��X�т̔��́A�y�і�U�z�̉e�����傫�������B�M�t�`���E�A�q���M�t�`���E�A�I�I�����T�L�̃`���E�ނł́A����ɉ����ė��l���傫�Ȉ��͂ƂȂ��Ă���B���l�ɂ��e�����傫����Ƃ��Ă͑��ɃQ���W�{�^�����������B
�@�S�̓I�ɂ݂�ƁA�M�t�`���E�ƃ^�K���ɑ���e��v���̉e�����傫���A�I�I�����T�L�A�Q���W�{�^���A�n�b�`���E�g���{������Ɏ����ő傫�ȉe�����Ă���B
�@
�@������@���͎w�W�����Ɠ���ł��邪�A�����Ώێ�͗\�ߒ�߂��Ă��炸�A�e�s���{���ŁA���L�̊�ɂ��A50�`100���I�肵�āA���̐����n�E���E�����̌���Ȃǂ������B
A�@���{�ł͂����ɂ����Y���Ȃ���
B�@���z�悪���肳��Ă����
C�@���z���E�Ǝv�����
D�@��ł̂�����̂����
E�@��ł����Ǝv�����
F�@���l�ɂ�茸���̂͂�������
G�@���w�W�ƂȂ蓾���
����ɁA����̕��ތQ�ɕ邱�Ƃ�����邽�߁A�ł��邾�������̖ڂɂ킽��悤�ɑI�肵�A���̌���22��1,754��̍����ނ��I�ꂽ�B
�����I���ʂɐ����������̂��\�Q�|�V�|�V�ł���B
�I���ʂ̎퐔�ɂ��Ă݂�ƁAB���ł�����1009��i�y�ш���j�œ��荩���S�̂̂U���߂������̊�ɊY������B�����ő����̂�699��i��S���j��C�ŁA�ȉ�D�i313��j,A�i273��j,G�i195��j,E�i65��j,F�i61��j�̏��ł������B
�@
�U�D�I���ɂ��ی�̌��n����݂����荩���ނ̈ʒu�Â�
�@�I����G�������ƁA���z��\�����́iA�`C�j�Ɛ����̌X���i���ɐ�ŁA�����Ɋւ���j��\�����́iD�`F�j�Ƃɕ�������B
���҂́u�ی�̕K�v���v�Ƃ����ϓ_���爽�镪�ތQ�����ŗD�揇�ʂ�t���悤�Ƃ���ہA�l�����ׂ��ł���A�������I�ɔ���\�Ȋ�ł���B
�A�D�I��A�́u���{�ł͂����ɂ����Y���Ȃ��Ǝv�����v�ƒ�`����邪�A����ɂ͓�̏ꍇ���z�肳���B���Ȃ킿�P���E�I�ɂ݂Ă����{�ɂ����Y���Ȃ����́i���{���Y��i���͈���j�j�Q���E�I�Ɍ���Α��̒n��i��Ƃ��ăA�W�A�n��j�ɂ��Y���邪���{�ł͗B��̒n��ɂ����Y���Ȃ����́A�̓�ł���B
���u�B��̎Y�n�v�Ƃ݂Ȃ��ׂ��n��̍L���肪���m�łȂ����A�Ƃ�܂Ƃ߂ɓ����������ψ��̋��ʓI�F���Ƃ��ẮA�s�A���ȕ��z������ꍇ�ɂ́A���͈̔͂������ނ˓s�{�����x�A�A���I�ȕ��z������ꍇ�ɂ͐����ɂ܂����邱�Ƃ����邪�A���̏ꍇ�ł��ʐϓI�ɂ͂P�s�{�����x�̂��́A�Ƃ����Ƃ���ł���B���������āA�k�C���S��ɍL�����z������̂𑼒n��Ɍ��o���Ȃ�����Ƃ����ākA�l�Ɋ܂߂�̂͑Ó��ł͂Ȃ��B
�C�D�I��B�́u���z�悪������̒n��Ɍ��肳��Ă����v�ƒ�`�������̂ł��邪�A���̏ꍇ�ɂ��A�̂P�Q�̋敪����������B�u��̒n��v�Ƃ́A�����ނ˂P�n�����x�ł���A�S���I�ɕ��z����ꍇ�͂T�`�U�����Ƃ���̂��Ó��Ǝv����B
�E�D�I��C�́u���ʎ�ł����Ă��k���E����ȂǕ��z���E�ɂȂ�Ǝv����Y�n�ɕ��z�����v�ł���B���̊�ɂ����Ă��P�Q�̃J�e�S���[�敪�͐�������Ǝv���邪�A��Ƃ��ĂQ�̃J�e�S���[�ɑ����A�嗤�̖k���n��̂��̂������A����A�W�A�n��̂��̂��k����ƂȂ�ꍇ�������悤�ł���B
�i�Q�j�����i��ŁA�����X���j��\���I��
�A�D�I��D�́u���Y�n��ɂ����Đ�ł̊�@�ɕp���Ă����v�ł���A��̓I�Ȕ��荪���͖��m�łȂ����A�n��I�Ȑ�ł̑O�i�K�Ɖ��߂����B
�C�D�I��E�́u�ߔN���Y�n��ɂ����Đ�ł����ƍl�������v�ł���A�����̕����\���Ƃ͕ʂɁA�����_�ł͓��Y�n��ł͐������m�F����Ȃ��Ȃ�����ł���B
�E�D�I��D�́u�Ǝ҂��邢�̓}�j�A�Ȃǂ̗��l�ɂ��A���Y�n��ł̌̐��̒������������S�z������v�ŁA����͐����̐������������v���̂����ł���߂đI��I�ł���i���̈��͂͂͂��߂���킻�̂��̂Ɍ������Ă�����̂ł͂Ȃ��j�A�������̕ی�̓_����͖����ł��Ȃ��̏W�ɂ��Ď�グ�����̂ŁA�����́AD���x���邢�͂��̑O�i�K�Ƃ݂Ȃ�����B
���z��\���I���ɂ́u��i���͈���j���̂��̂̐�ł̊댯���̒��x�v���邢�́u�i���Y�n��ɂ����Đ�ł����ꍇ�́j�����̉\���̒��x�v����݂������N�t���Ƃ����܈ӂ����邱�Ƃ͖����ł���B���Ȃ킿�A���{���Y��̗B��̐����n�ŁA���̌̌Q����ł���A���Y��͑S���̉\���͂Ȃ��u�킻�̂��̂���Łv�������ƂɂȂ�B���ӑ嗤���ɓ���킪���݂���ꍇ�́A�l�דI�ȓ�����ʂƂ���A�����̉\���͂قƂ�ǂȂ��Ƃ����Ă悢���낤�B���z�悪���肳����̏ꍇ�́A���������ۂ���Ă��邩�A���������Ύ��ӂ���̐N�����ɂ���ĉ���\���͏\�����邾�낤�B�������A���z�̕s�A���������������̂͂��̌���łȂ��B
���z�̌��E�n��́A���Y��̕K�v�Ƃ��鐶�����������e���x�����ς��ł��邱�Ƃ��������߁A�e�Ղɒn��̌Q�̐�ł������Ղ����A����͎�̕��z��̏k���������炷���A�킻�̂��̂̑������������ɂ͎���Ȃ����낤�B
���������āA�����Ɛ�����\���D�EE�EF��g������A��̕ۑ��Ƃ����ϓ_����J�����ׂ����ʂ����̂����疾�炩�ƂȂ�B�i�\�Q�|�V�|�W�j
��L�̌����܂��A���荩����1754������̗v�̂Ń��X�g�A�b�v�����i�����ҁj�B
�A�D�I��A
�I��A�ɊY�������́A���݂̂Ƃ���A��ŁA�����̒��F�߂��Ȃ�����܂߂đS�탊�X�g�A�b�v�����B
�C�D�I��E
�S�탊�X�g�A�b�v�����B
�E�D�I��B�ł���A����D�ł������
�I��B�y��C�ɂ��ẮA���ނ��\���i��ł��Ȃ��O���[�v�⏬�^��A������̑����O���[�v�ł͕��z�悪�c������Ă��炸�A���̊�̂��Ă͂߂͍���ł��邱�Ƃ������B�����ŕ��ނ��悭�i��ł���ڂɂ��Ղ���^��̑����O���[�v�Ɍ����ă��X�g�A�b�v��i�߂��B�ΏۂƂ����̂́A��x�ځA�����ځA�����ځi�J�}�L���ށj�A�����ځi�������ځj�A�����ځi���ށj�A�����ځi�n���~���E�ȁA�I�T���V�ȁA�S�~���V�ȁA�Q���S���E�ȁA�N���K�^���V�ȁA�R�K�l���V�ȁA�^�}���V�ȁA�J�~�L�����V�ȁj�ł���B����ɊY������͍̂��v�{�{�{��ł������B
�G�D�����̓s���{����D����F�Ƃ��đI�肳�ꂽ���́i�T���ȏ�j
�I�D�I��G
�I��G�́u���w�W�Ƃ��ēK���ł���ƍl�������v�ł���A���̓_�ɂ��Ă͍���̌����ۑ�Ƃ���B
�@
���鐶���̑��݂̗L���ɂ���āA�n����̓���̊��̔c�����s�����Ƃ���ꍇ�ɂ͓��Y�n����ɂ����Ď��F���������{����Ȃ���Ȃ�Ȃ��B�w�W�����ނ̒����͌���ꂽ���̒������ɂ����̂ł��������߁A���������̎��W�E���������S�ƂȂ炴����A���n�m�F�╷���݂́A�\�Ȕ͈͂ŕ�[�I�Ɏ��{���ꂽ�ɂ����Ȃ��B���������ă^�K���̂悤�ɈȑO�͂�������ӂꂽ��ł��������߁A�����҂̊S�������M��������L�^���S�̂ɖR�����Ƃ������������ŁA�{�����̌��ʂ��s���{���̌����̕��z�𐳊m�ɔ��f���Ă���Ƃ�����Ԃɂ͎����Ă��Ȃ��̂�����ł���B
�������A�n��I�ɂ݂�Ώ��̌����͂�����x�F�߂�ꂽ�Ƃ��Ă��A�{�����͓��Y�����ނɊւ���S���I�ȕ��z�����Ƃ��Ă͖��]�L�̂��̂ł���A���̌��ʂ͌����_�ł͕��z��c��������B�ꂩ�ő�̏��ʂ�L���鎑���ł���B
�����ŁA�����w�W�����ނ̕��z�A�������̏��I�Ȋ��w�W�Ƃ��Ă̒n������̔c���́A�s���{�����x���ł͏\���ȍ����������Ɣ��f���A��q�̍�Ƃ��s�������̂ł���B
���̍�Ƃɂ���ĂȂ��ꂽ�s���{�����Ƃ̊��c���́A���n�m�F��������x�s�Ȃ�ꂽ�Ƃ���ł͂��ꂼ��̒n��̒����҂��L�q���Ă���n��̏Ƃ����ނˈ�v���Ă���Ǝv��ꂽ�B
���������łɏq�ׂ��悤�ɑS���ɍL�����z���邱�Ƃ������Ƃ��đI�肳�ꂽ�w�W�����ނł͂��邪�A���R���z���Ȃ��n��������ΏۂƂ��鎩�R����ԗ��ł��Ȃ��ꍇ��A���z���Ă��Ă����z��̌��E�t�߂̂��͎̂��R��Ԃł��Ǝ�ł��邽�߂��A�l�דI�ȉe���ȏ�ɏ�Ԃ����Ă���ƍl������ꍇ���������B
���̓_�����P���邽�߂ɂ́A���̂悤�Ȓn��ł͂���ɑւ����I�肷�邱�Ƃ��ώ��ȏ��ʂ̑���ƂƂ��ɍ���̉ۑ�ƂȂ邾�낤�B
���w�W�����Ƃ������i��A�S���I�ɍL�����z���A���A���Ȃ蕁�ʂɐ������Ă���i�͂��́j�킪�A�����ΏۂƂ��đI�肳�ꂽ���A�����̌��ʁA�����̎�̑����́A�����n�̔j��≘���A���l���ɂ��A���������������Ă��邱�Ƃ����炩�ƂȂ����B���ɐl�Ԋ����̐���ȕ��n���̏��~���i�r���⎼�n�j����Ƃ����ށi�^�K����n�b�`���E�g���{�j�ɂ��̌X���������ł������B
����̊��݂��đI�肳�ꂽ���荩���ނ́A1754��Ƃ��������ɏ�����B�����̎�ɑ��āA�u��v�̕ۑ��|��ł̊댯���Ƃ����ϓ_����Č������������B
�@